Biliary fibrosis is an important but neglected pathological feature in hepatobiliary disorders: from molecular mechanisms to clinical implications
-
Jinyu Zhao
 ,
Jinqiu Yuan
,
Yanyan Lin
,
Hans J. Schlitt
and
Wenbo Meng
,
Jinqiu Yuan
,
Yanyan Lin
,
Hans J. Schlitt
and
Wenbo Meng


Abstract
Fibrosis resulting from pathological repair secondary to recurrent or persistent tissue damage often leads to organ failure and mortality. Biliary fibrosis is a crucial but easily neglected pathological feature in hepatobiliary disorders, which may promote the development and progression of benign and malignant biliary diseases through pathological healing mechanisms secondary to biliary tract injuries. Elucidating the etiology and pathogenesis of biliary fibrosis is beneficial to the prevention and treatment of biliary diseases. In this review, we emphasized the importance of biliary fibrosis in cholangiopathies and summarized the clinical manifestations, epidemiology, and aberrant cellular composition involving the biliary ductules, cholangiocytes, immune system, fibroblasts, and the microbiome. We also focused on pivotal signaling pathways and offered insights into ongoing clinical trials and proposing a strategic approach for managing biliary fibrosis-related cholangiopathies. This review will offer a comprehensive perspective on biliary fibrosis and provide an important reference for future mechanism research and innovative therapy to prevent or reverse fibrosis.
Introduction
Fibrosis, a pathophysiological repair response to tissue injury, is often characterized by the inflammatory-mediated accumulation of extracellular matrix (ECM) components such as collagen, leading to scar formation [1, 2]. In physiology, fibrous connective tissue maintains organ structure and function, replacing necrotic or damaged cells during injury. However, pathological fibrosis arises from repeated or sustained tissue injury and a dysregulated balance between collagen production and degradation by myofibroblasts, resulting in organ sclerosis and dysfunction [3]. Approximately one-third of global deaths are attributed to organ failure from fibrosis [4]. Fibrosis affects nearly all organs and is implicated in chronic inflammatory disorders, autoimmune conditions, and neoplasms, notably in the skin, lung [5], heart [6], liver [7], kidney [8], pancreas [9], bone marrow [10], and intestine [11, 12] (Figure 1). Research data indicates an annual incidence of fibrosis-related diseases as high as 4,968 cases per 100,000 person-years, with a rising trend due to increasing awareness [13].
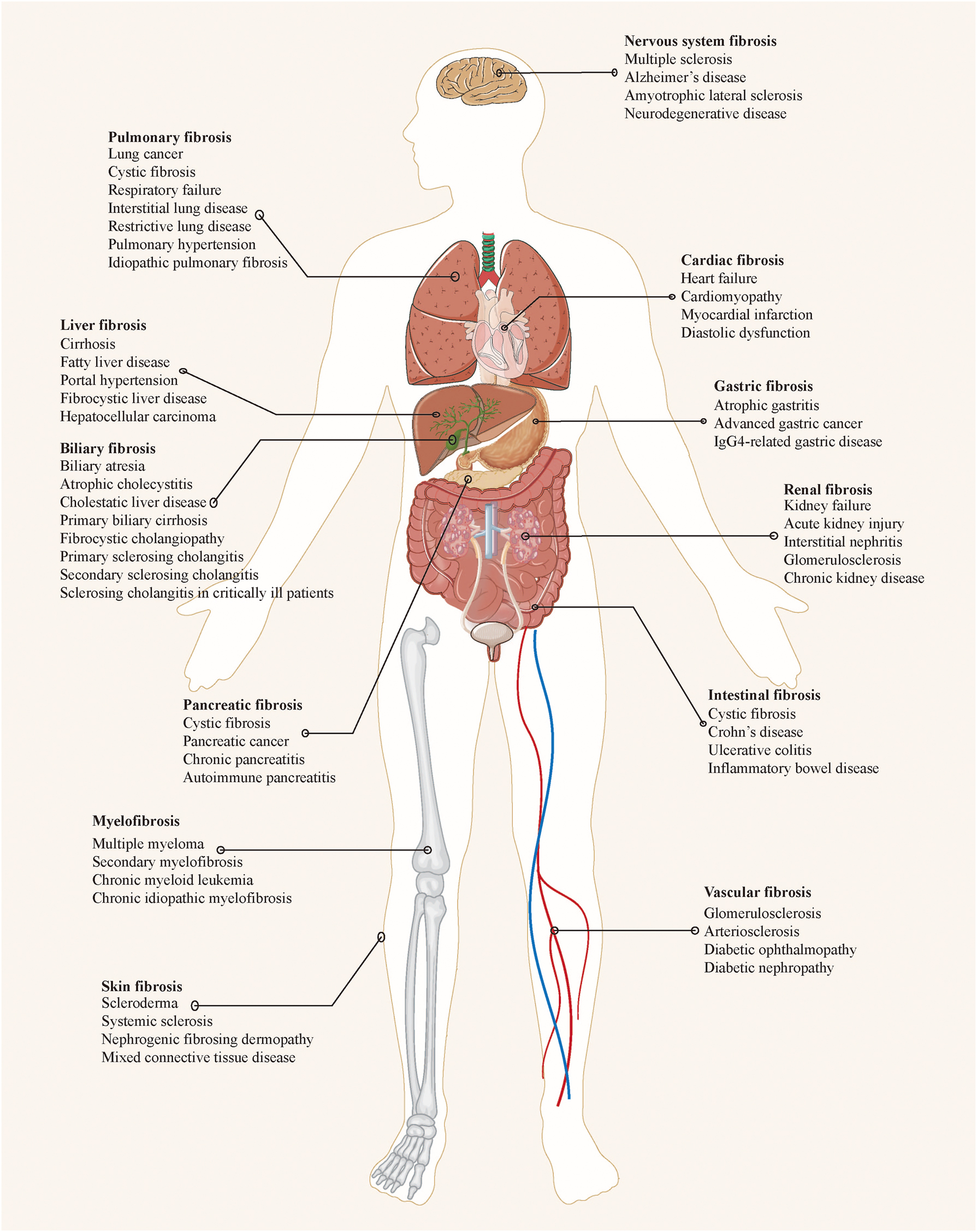
Common organ fibrosis and related diseases.
Biliary fibrosis secondary to repeated or prolonged biliary epithelium injury and inflammation has been identified as an important but neglected pathological feature in specific cholangiopathies such as biliary atresia (BA) [14], primary sclerosing cholangitis (PSC) [15], secondary sclerosing cholangitis (SSC) [16], primary biliary cholangitis (PBC) [17], and IgG4-related sclerosing cholangitis (IgG4-SC) [18]. Biliary fibrosis may promote the silent development and progression of both benign and malignant biliary diseases through pathological repair mechanisms secondary to biliary injury. Elucidating the relevant etiology and pathogenesis is helpful to the inhibition or reversal of biliary fibrosis, so as to aid the clinical management of cholangiopathies.
This review highlighted the importance of biliary fibrosis in hepatobiliary disorders and detailed relevant etiology and pathogenesis. We also summarized important signaling pathways involved in biliary fibrosis and potential therapeutic strategies for fibrosis-related cholangiopathy. The primary objective of this review is to provide a reference point to contribute to further mechanism research, drug development, and clinical trials of biliary fibrosis.
Definition and clinical features of biliary fibrosis
Definition and scope of biliary fibrosis
Biliary fibrosis refers to the diffuse excessive deposition and abnormal distribution of ECM (e.g., collagen, glycoprotein, proteoglycan, etc.) caused by the pathological repair response of biliary system secondary to chronic injury [19], which is closely related to the progress and prognosis of various cholangiopathies. Biliary fibrosis manifests as sclerosis, wall thickening and luminal stenosis of bile duct, or sclerosis and atrophy of gallbladder. Although both biliary fibrosis and hepatic fibrosis may progress to cirrhosis, there are significant differences in etiology, clinical characteristics, pathology, etc. [20, 21] (Table 1).
Differences and connections of biliary and hepatic fibrosis.
| Characteristic | Biliary fibrosis | Hepatic fibrosis |
|---|---|---|
| Cause | Cholangiocyte injury | Hepatocyte damage |
| Related disease | Biliary atresia, SSC, biliary parasitosis, cholestatic liver disease such as PBC, PSC, IgG4-SC, etc. | Chronic viral hepatitis, chronic alcoholic hepatitis, NASH, AIH, etc. |
| Pathogenesis | Chronic biliary inflammation; Reactive duct cells activate myofibroblasts; ECM accumulates around portal veins; PFB is the main source of myofibroblasts. |
Chronic liver inflammation; Hepatocytes activate myofibroblasts; ECM accumulates around centrilobular veins; HSC is the main source of myofibroblasts. |
| Fibrosis pattern | Portal-to-portal evolution | Portocentral or pericentral evolution |
| Clinical symptoms | Fatigue, pruritus, abdominal pain, jaundice, etc. | Fatigue, loss of appetite and weight, portal hypertension symptoms, etc. |
| Laboratory indicators | ALP, γ-GT, ALT, AST, bilirubin, ANAs, AMAs, IgG4, etc. | ALT, AST, ALB, etc. |
| Imaging features | Abnormalities of biliary system, mainly including sclerosis, stricture, occlusion or absence of biliary tree. | Abnormalities of liver parenchyma, mainly including increased elasticity, atrophy, and nodule of liver. |
| Progression | Relatively rapid progression to biliary cirrhosis (the liver gradually becomes larger). | Relatively slow progression to hepatic cirrhosis (the liver gradually becomes smaller). |
| Connections | Biliary fibrosis is usually accompanied by obvious cholestasis, and excessive intrahepatic cholestasis can cause hepatocyte damage. Therefore, hepatic fibrosis can occur in advanced stages of biliary fibrosis. Liver transplant is an effective treatment at the end stage. | |
-
SSC, secondary sclerosing cholangitis; PBC, primary biliary cholangitis; PSC, primary sclerosing cholangitis; IgG4-SC, IgG4-related sclerosing cholangitis; NASH, nonalcoholic steatohepatitis; AIH, autoimmune hepatitis; ECM, extracellular matrix; PFB, portal fibroblast; HSC, hepatic stellate cell; ALP, alkaline phosphatase; γ-GT, γ-glutamyl transpeptidase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; ANAs, anti-nuclear autoantibodies; AMAs, anti-mitochondrial antibodies; ALB, albumin.
Biliary fibrosis is a common pathological feature in cholangiopathies, including BA [22, 23], PBC [17, 24, 25], PSC [17, 26], [27], [28], IgG4-SC [17, 29], and SSC [30], [31], [32] (Table 2). SSC is a general term for sclerosing cholangitis with definite etiologies such as chronic biliary obstruction, infection, immunity, ischemia, toxin injury, mechanical damage, genetics, etc. [33]. The main biliary disorders of SSC include sclerosing cholangitis in critically ill patients (SC-CIP), AIDS cholangiopathy, ischemic cholangiopathy, recurrent suppurative cholangitis, drug-related cholangitis, hereditary cholestasis and biliary parasitic disease. Notably, biliary fibrosis in cholangiopathies may eventually lead to serious outcomes such as biliary obstruction, biliary cirrhosis, biliary malignancy, and secondary liver failure without effective treatment.
Evidence of biliary fibrosis in cholangiopathies.
| Cholangiopathy | Pathological features | Imaging features | Serum indicators |
|---|---|---|---|
| BA [22, 23] | Bile duct inflammation; Progressive biliary fibrosis; Ductular reaction; Bile duct loss; Bile duct plate malformations. |
Triangular cord sign; Liver subcapsular vascular sign; Gallbladder ghost triad; Bile duct disappearance; Increased liver elasticity. |
MMP7; γ-GT; DBIL. |
| PBC [17, 24, 25] | Chronic non-suppurative destructive cholangitis; Florid bile duct lesions; Ductular reaction; Bridging fibrosis; Biliary cirrhosis. |
Liver diffuse enlargement; Liver nodule formation; Liver parenchymal heterogeneity; periportal halo sign; Irregular bile duct structure. |
ALP; γ-GT; ALT; AST; DBIL; AMAs; ANAs; IgM. |
| PSC [17, 26], [27], [28, 501] | Chronic periportal inflammation; Onion skin periductal fibrosis; Ductular reaction; Bridging fibrosis; Biliary cirrhosis. |
Band-like stricture of bile ducts; Beaded appearance of bile ducts; Pruned-tree appearance of biliary tree; Diverticulum-like dilation of bile duct. |
ALP; γ-GT; ALT; AST; DBIL. |
| IgG4-SC [17, 29] | Lymphocyte and plasma cell infiltration; Bile duct storiform fibrosis; Obliterative phlebitis. |
Isolated or continuous bile duct stricture; Concentric thickening of bile duct wall; Withered-tree appearance of biliary tree; With extrahepatic diseases like AIP, IBD. |
IgG4; ALP; γ-GGT; ALT; AST. |
| SSC (SC-CIP) [30], [31], [32], [33] | Periportal inflammation; Peribiliary fibroplasia; Ductular reaction; Progressive fibrosis of intrahepatic bile ducts. |
Multiple irregular strictures of bile ducts; Tube-shaped structures in bile ducts; Beaded appearance of bile ducts; Pruned-tree appearance of biliary tree. |
ALP; γ-GT; ALT; AST; TBIL. |
| Atrophic cholecystitis | Chronic inflammation; Gallbladder wall fibrous thickening; Scar formation. |
Gallbladder shrinkage; Diffuse thickening of gallbladder wall; Disappearance of gallbladder. |
Unclear |
-
BA, biliary atresia; MMP7, matrix metalloproteinase 7; γ-GT, γ-glutamyltranspeptidase; DBIL, direct bilirubin; PBC, primary biliary cholangitis; ALP, alkaline phosphatase; ALT, aspartate aminotransferase; AST, aspartate aminotransferase; AMA, anti-mitochondrial antibodies; ANAs, anti-nuclear autoantibodies; PSC, primary sclerosing cholangitis; IgG4-SC, IgG4-related sclerosing cholangitis; SSC, secondary sclerosing cholangitis; SC-CIP, sclerosing cholangitis in critically ill patients; TBIL, total bilirubin.
Epidemiology of biliary fibrosis
The overall incidence and prevalence of biliary fibrosis remain unknown, but epidemiological data on fibrosis-related cholangiopathies in the literature provide a reference. For PBC, the estimated global incidence and prevalence are 17.6 per million and 146 per million, respectively [34]. For PSC, Nordic population studies suggest a prevalence of 0.15 per million for small duct PSC and 0.91–1.30 per million for classical PSC [26]. For BA, the global incidence is approximately 0.55–1.3 per 10,000 live births [22]. For IgG4-SC, the estimated overall prevalence based on autoimmune pancreatitis is 0.2 per million [29]. Moreover, 6 %–35 % of the patients develop biliary complications such as anastomotic or non-anastomotic strictures after liver transplantation [35], which are considered a major cause of morbidity, graft failure, and mortality after liver transplantation [36]. However, there are obvious regional differences in the epidemiology [34], which is not only affected by the regional distribution of pathogenic factors, but also related to the cognition of the disease. Meanwhile, the prevalence of cholangiopathy also varies with age and gender, and the role of sex hormones in biliary fibrosis is worthy of attention.
Clinical manifestations of biliary fibrosis
Biliary fibrosis is insidious and lacks clinical manifestations. In the early stages of the disease, the main manifestations are abnormalities in molecular biological indicators, including alkaline phosphatase (ALP), γ-glutamyl transpeptidase (γ-GT), alanine aminotransferase (ALT), aspartate aminotransferase (AST), serum bilirubin, autoantibodies and IgG4. These indicators serve as reflections of biliary injury and inflammation, altered liver function, and bile acid metabolism, guiding the initial screening and severity assessment of biliary fibrosis. Fatigue, itching, abdominal pain and jaundice are the predominant but nonspecific symptoms, with portal hypertension occurring in the advanced stages [37], [38], [39].
Imaging features of biliary fibrosis
The foremost and definitive indication of biliary fibrosis manifests as occlusion and loss of bile ductules, fibrous thickening and stenosis of large bile ducts, and sclerosis and shrinkage of the gallbladder. Classical non-invasive imaging methods such as ultrasound (US), computed tomography (CT) and magnetic resonance cholangiopancreatography (MRCP), are currently not used as a standard for a definite diagnosis of biliary fibrosis, but guide initial screening and exclusionary diagnosis. Fibrous stenosis of bile ducts is one of the important imaging findings of biliary fibrosis [17, 40]. In PSC, multifocal stenosis of intrahepatic ducts (15 %–25 %) or extrahepatic ducts (5 %–10 %) with mildly dilated segments manifests as “beaded” changes in the biliary tract imaging findings [40, 41]. In PBC, periportal fibrous component deposition and hepatocyte parenchymal regression form a “periportal halo” sign on MRI [17, 42, 43]. In IgG4-SC, its characteristic imaging changes are segmental and long (confluent) strictures with pre-stenotic dilation [16, 40]. In ischemic SSC, defects in the intrahepatic biliary tree are observed in the early stages, and diffuse intrahepatic bile duct strictures are observed in the advanced stages [16]. In addition, invasive imaging methods such as endoscopic retrograde cholangiopancreatography (ERCP), endoscopic ultrasound (EUS), intraductal ultrasound (IDUS), per-oral cholangioscopy (POC) and endoscopic ultrasound fine needle biopsies (EUS-FNB), allow directly endoscopic observation and biopsy of the biliary tract [44, 45], aiding in the definitive diagnosis of biliary fibrosis.
Pathology of biliary fibrosis
Biliary fibrosis is essentially an excessive repair response to biliary tissue damage, which can occur anywhere in the biliary system, including intrahepatic bile ducts, extrahepatic bile ducts, and gallbladder. Macroscopically, large bile duct fibrosis is characterized by sclerosis, wall thickening and lumen stenosis of bile ducts, with or without bile duct dilatation; small bile duct fibrosis is characterized by occlusion or loss of bile ducts, and secondary biliary cirrhosis; gallbladder fibrosis is characterized by sclerosis, thickening and atrophy of gallbladder, and gallstone formation. Microscopically, the histological characteristics of biliary fibrosis include chronic biliary tract inflammation, bile duct reaction, infiltration, and sustained activation of myofibroblasts, excessive ECM (e.g., collagen fibers, elastic fibers, fibronectin and laminin, etc.) accumulation in bile duct interstitium and basement membrane, fibrous thickening and stenosis of bile ducts, and occlusion and loss of small intrahepatic bile ducts. For instance, “florid duct lesion” of PBC [24, 46], “onion skin” of classical PSC [28, 47], and “storiform fibrosis” of IgG4-SC [17, 29] are pathological markers of biliary fibrosis.
Diagnosis of biliary fibrosis
The definitive diagnosis of biliary fibrosis currently depends on histopathology. ERCP, POC, and EUS-FNB are the main methods of biliary tract biopsy [44, 45, 48]. Liver biochemical indicators (i.e., ALP, γ-GT, ALT, AST, bilirubin) and non-invasive imaging are used for initial screening of biliary fibrosis. However, there is a lack of established criteria for diagnosing and grading of biliary fibrosis. The combined application of endoscopy and imaging has a good prospect in the evaluation and diagnosis. Elastography of US has been used to assess severity of fibrosis in PBC by measuring duct wall stiffness. EUS significantly enhances the diagnostic capabilities of both endoscopy and US [49]. Contrast-enhanced EUS (CE-EUS) directly observes the perfusion features of the biliary tract [50] and has been proven to accurately identify various gallbladder diseases, including cancer, polyps, stones, and inflammatory hyperplasia. ERCP-IDUS shows advantages in the differential diagnosis of benign and malignant biliary strictures [51, 52]. Specific serum molecular markers from cholangiopathies characterized by biliary fibrosis such as IgG-4, anti-mitochondrial antibody, anti-nuclear autoantibodies, matrix metalloproteinase 7, etc. may also aid in the diagnosis of biliary fibrosis. Notably, the enhanced liver fibrosis (ELF) score, an algorithm based on serum concentrations of procollagen-III aminoterminal propeptide, tissue inhibitor of matrix metalloproteinase-1 and hyaluronic acid, has been confirmed to reflect active fibrosis and the extent of disease in IgG4-SC [53], PBC [54] and PSC [55]. However, the role of these specific molecular markers of fibrosis in bile is unknown. As a side note, the role of intestinal and biliary flora in the diagnosis of biliary fibrosis is also worthy of attention.
Etiology of biliary fibrosis
Repeated or prolonged biliary epithelial cell injury and chronic inflammation underlie the pathogenesis of biliary fibrosis. Factors related to biliary tract injury may contribute to the development and progression of biliary fibrosis, including genetics, immunology, infection, ischemia, cholestasis, and iatrogenic injury (Table 3).
Predominant etiology and related pathogenesis of biliary fibrosis.
| Etiology | Related cholangiopathies | Pathogenesis |
|---|---|---|
| Hereditary | PBC; PSC; BA; Pancreaticobiliary maljunction; Hereditary cholestasis diseases (e.g., PFIC, BRIC, CBASD, and alagille syndrome); Cystic fibrosis-associated cholangiopathy. |
1) Abnormal development of biliary tract; 2) Cholangiocyte function defects; 3) Bile acid transport disorder; 4) Epigenetic effects. |
| Immune-mediated | PBC; PSC; IgG4-SC; Eosinophilic cholangitis; Mast cell cholangiopathy; Post-liver transplant cholangiopathy. |
1) Immunologically induced biliary epithelial injury; 2) Immunologically induced inflammation; 3) Hepatic allograft rejection. |
| Infectious | Recurrent suppurative cholangitis; Biliary parasitosis (e.g., liver fluke, ascaris); AIDS-related cholangiopathy (e.g., cryptosporidiosis, microsporidiosis). |
1) Chronic inflammation of biliary mucosa; 2) Direct injury to biliary epithelium. |
| Ischemic | Post-liver transplant cholangiopathy; Ischemia-related SSC; SC-CIP. |
Impairment of arterial blood flow, at the level of the major hepatic artery branch or at the peribiliary vascular plexus. |
| Iatrogenic | COVID-19-related cholangiopathy; Surgery-related SSC; Drug-related SSC; SC-CIP. |
1) Drug-induced biliary epithelial injury; 2) Direct injury to biliary epithelium by operation; 3) Insufficient blood and oxygen supply to cholangiocytes; 4) Systemic inflammatory response syndrome. |
| Cholestatic | Cholestatic liver disease (e.g., PBC, PSC, IgG4-SC, SSC, SC-CIP, BA, PFIC, BRIC, CBASD) | 1) Toxic bile-induced cholangiocyte injury; 2) Microflora-induced biliary inflammation. |
-
PBC, primary biliary cholangitis; PSC, primary sclerosing cholangitis; IgG4-SC, IgG4-related sclerosing cholangitis; BA, biliary atresia; PFIC, progressive familial intrahepatic cholestasis; BRIC, benign recurrent intrahepatic cholestasis; CBASD, congenital bile acid synthesis disorders; SSC, secondary sclerosing cholangitis; SC-CIP, sclerosing cholangitis in critically ill patients.
Genetic factors
Genes regulate the normal development of the biliary tree, and interactions between biliary epithelial and mesenchymal tissues achieve precise regulation of the formation and remodelling of the biliary system. Familial and genetic studies have highlighted the importance of genetic susceptibility in fibrosis of cholangiopathies, including PBC [56], PSC [57], BA [58, 59], pancreaticobiliary maljunction [60, 61], choledochal cysts [62], fibrocystic liver disease [32, 63], and cystic fibrosis [64, 65]. Genome-wide association studies have identified several alleles associated with PBC susceptibility at human leukocyte antigen (HLA) and non-HLA loci [24, 66, 67], including HLA-DRB1*11 [68], HLA-DRB1*13 [68], HLA-DQB1*06:04 [68], HLA-DQB1*03:01 [68, 69], HLA-DRB1*08:03 [68], HLA-DPB1*17:01 [68], and IL-12 pathway [24, 66, 70]. Comprehensive research on hereditary cholestasis diseases has confirmed the genetic loci associated with specific disorders [71], such as ATPB8B1, ABCB11, ABCB4, TJP2, NR1H4 and MYO5B of progressive familial intrahepatic cholestasis (PFIC); ATP8B1 and ABCB11 of benign recurrent intrahepatic cholestasis (BRIC); HSD3B7, AKR1D1, CYP7B1 and AMACR of inborn errors of bile acid synthesis (IEBAS); JAG1 and NOTCH2 of Alagille syndrome. In addition, various mouse models of biliary fibrosis have been successfully established through gene knockout methods [56].
Immune factors
Immune dysregulation is commonly observed in fibrosis-related cholangiopathies such as PBC and PSC. The innate and adaptive immunity are involved in the pathogenesis of PBC, as indicated by the presence of granulomatous inflammation, the hypersecretion of proinflammatory cytokines and IgM, the elevation of NK and NKT cells, as well as the increase of antigen-specific CD4+ and CD8+ T cells [24]. CD8+ T cells, as the predominant infiltrating lymphocytes in the liver tissues of PBC patients, promote the injury and apoptosis of cholangiocytes by expressing FasL and secreting perforin. Immune cells (e.g., macrophages, eosinophils and T cells) play roles in cholangiocyte activation and peribiliary inflammation in PSC, thereby promoting biliary fibrosis [72]. Unconventional T cells that are important for recognition of bacterial pathogens also play a role in PSC and tend to localize to areas of fibrosis [73, 74]. In addition, autoimmune antibodies of IgG4-SC, PBC and PSC [24, 29, 71], elicit an autoimmune response, instigating progressive biliary damage and inflammation, and promoting the development and progression of biliary fibrosis.
Infectious factors
Biliary infection is a common cause of biliary epithelial injury, including biliary parasitosis, biliary and intestinal flora dysbiosis, and viral infection. Parasites induce persistent biliary inflammation and immune activation, primarily through both direct physical damage by the worms and chemical stimulation via excretory metabolites within the biliary tract. Studies have indicated a strong association between biliary fibrosis [75, 76] and the parasites Clonorchis sinensis (also known as liver fluke) [76, 77] and Ascaris lumbricoides [78]. Chronic inflammation of the biliary mucosa due to microbial-mucosal interactions is another pathway for biliary tract injury, with retrograde biliary infection by gut microbes as a source of biliary microbes. Biliary Helicobacter pylori [79] and Enterococcus faecalis [80] are significantly linked to the development and progression of biliary fibrosis. Alterations in intestinal flora are also observed in biliary fibrosis, including reduced microbial diversity and increased proportions of specific flora such as Enterobacteriaceae, Enterococcus spp., and Verrucoccus spp. [80, 81]. Dysbiosis of intestinal flora may contribute to inflammation in the hepatobiliary system by influencing bile acid metabolism, intestinal permeability, utilization of short-chain fatty acids, and macromolecule metabolism[81]. Moreover, biliary fibrosis has been observed in patients infected with human immunodeficiency virus (HIV) [82], cytomegalovirus (CMV) [83], and coronavirus disease 2019 (COVID-19) [84, 85], but the exact mechanisms remain unclear.
Iatrogenic factors
The primary influence of iatrogenic factors on biliary fibrosis is biliary injury caused by drugs [16, 86] and surgery [87]. Pharmacological biliary injury primarily results in small bile duct epithelium damage and cholestasis. SSC has been reported after therapy with chemotherapeutics [88] (e.g., docetaxel [89], bevacizumab [90], 5-FU and leucovorin therapy [91], and also after using immune checkpoint inhibitors such as pembrolizumab and nivolumab [92], [93], [94], [95], [96], [97], other drugs such as ketamine [98] or methimazole [99], and even herbal supplements [100]. The principal surgical procedures associated with biliary tract injuries include cholecystectomy, bile tract exploration, and liver transplantation [101], [102], [103], [104], [105]. Surgical injuries to the bile duct encompass direct mechanical damage resulting from surgical manipulation and instrumentation, cholestasis associated with benign biliary stenosis following surgery, biliary ischemia from post-surgical vascularization and revascularization, and severe complications such as chronic pancreatitis and post-traumatic SSC [106, 107].
Biliary ischemia
The blood supply of the biliary system relies on various arteries, mainly including the branches of the proper hepatic artery, hepatic artery, and cystic artery [108]. Notably, the superior and middle sections of the common bile duct are vascularized by two principal arteries running along its lateral edges, known as the “parabiliary arteries”, making this region particularly susceptible to arterial supply-related complications [108]. The right and left hepatic ducts, along with the intrahepatic ducts, receive a dense nonaxial arterial network from small arteries branching off the right and left hepatic arteries, forming the peribiliary vascular plexus (PBP) [109]. The intimate anatomical connection between the PBP and cholangiocytes fosters intercommunication, and which plays a role in the regulation of normal cholangiocyte function, and it is linked to cholangiocyte dysfunction in cholangiopathies [108]. The cholangiocyte is highly vulnerable to ischemia, and although minor lesions lead to re-epithelialization, more severe injuries result in fibrosis and stricture formation [110]. Hepatic arterial ischemia also reduces the expression and function of hepatocellular bile transporters, resulting in cholestasis, localized or extensive damage to both the gallbladder and bile ducts [111, 112]. Common causes of biliary tract ischemia include [108]: (1) excessive interruption of the hepatic artery during operation; (2) surgical devascularization and injury to the blood vessels, such as liver transplantation, cholecystectomy, and bilioenteric anastomosis; (3) arterial obstruction induced by hepatic arterial infusion of chemotherapeutic drugs or interventional hemostasis [28]; and (4) underlying disease statuses predisposing to biliary ischemia, such as advanced acquired immune deficiency syndrome (AIDS) [109], post-traumatic shock [113], hereditary hemorrhagic telangiectasia [114, 115], polyarteritis nodosa [116, 117], and atherosclerosis [109, 118]. The associations between systemic diseases and biliary ischemia have also been reported, such as sickle cell anemia [119], Henoch-Schönlein purpura (HSP) [120], systemic lupus erythematosus (SLE) [121], antiphospholipid syndrome (APS) [122], and paroxysmal nocturnal hemoglobinuria (PNH) [123]. Moreover, post-liver transplantation cholangiopathies are associated with severe damage to peribiliary gland (PBG) [124, 125], manifesting as poor regeneration of the biliary epithelium due to the destruction of progenitor cells and insufficient blood supply [126, 127].
Cholestasis
Bile contains diverse components including bile acids, electrolytes, lipids, proteins, and both endogenous and exogenous compounds. Bile secretion and transport disorders lead to retention of bile in the liver and blood, resulting in intrahepatic or extrahepatic cholestasis. Cholestasis serves as a mediator for biliary epithelial injury induced by various profibrotic factors, promoting the development and progression of biliary fibrosis. Bile acids, as a primary profibrotic factor in bile, induce biliary fibrosis through different mechanisms [81]: (1) direct toxic effects of bile salts on cholangiocytes; (2) continuous activation of myofibroblasts through bile acid signal pathways, stimulating the synthesis and excessive accumulation of ECM; and (3) the modification of intestinal flora by bile acid metabolites, promoting biliary injury and inflammation.
Pathogenesis of biliary fibrosis
General pathogenic mechanisms of organ fibrosis
Organ fibrosis involves multiple regulators and effectors, including cytokines, chemokines, and various cells. The development of organ fibrosis typically unfolds through four discernible stages [4]: (1) damage to specific endothelial and epithelial cells; (2) activation of effector cells, predominantly inflammatory cells and myofibroblast precursor cells; (3) synthesis and secretion of ECM; and (4) dynamic deposition and insufficient absorption of ECM. The deposition of collagen induces chronic hypoxia, further aggravating tissue damage, and perpetuating fibrosis in a vicious cycle. Chronic inflammation is a strong stimulant for fibrosis. However, once fibrosis is triggered, controlling inflammation alone proves inadequate to reverse the fibrosis process [128]. Moreover, biomechanical forces from collagen fibers promote myofibroblast activation, resulting in progressive ECM accumulation, tissue remodelling and fibrosis [2, 129]. Mechanical forces manifest as tensional stress, compressive stress, torsional stress, and shear stress [129], regulating gene expression and protein function related to fibrosis. For instance, biomechanical forces trigger myofibroblast activation and confer myofibroblast resistance to apoptosis through established mechanotransduction signaling pathways in fibrotic lung tissues [130]. Uniquely, fibrosis exhibits distinctive features across various organs [13], such as cardiac fibrosis [131], pulmonary fibrosis [132], renal fibrosis [133], liver fibrosis [7, 134], and multiorgan cystic fibrosis [135].
Cellular and molecular mechanisms of biliary fibrosis
Investigations have demonstrated that organ fibrosis is initiated within the “fibrotic niche”, characterized by intricate interactions between damaged parenchymal and nonparenchymal cells within the scarred region [3, 13, 136]. Single-cell sequencing studies examining cellular profiles in various fibrotic conditions, including idiopathic pulmonary fibrosis (IPF) [137], liver fibrosis [21], renal fibrosis [133], and systemic sclerosis, have unveiled the pivotal roles played by mesenchymal cells, immune cells, and distinct types of epithelial and endothelial cells in the fibrosis process. Here, we summarize in detail the cellular and molecular mechanisms of biliary fibrosis (Figure 2).
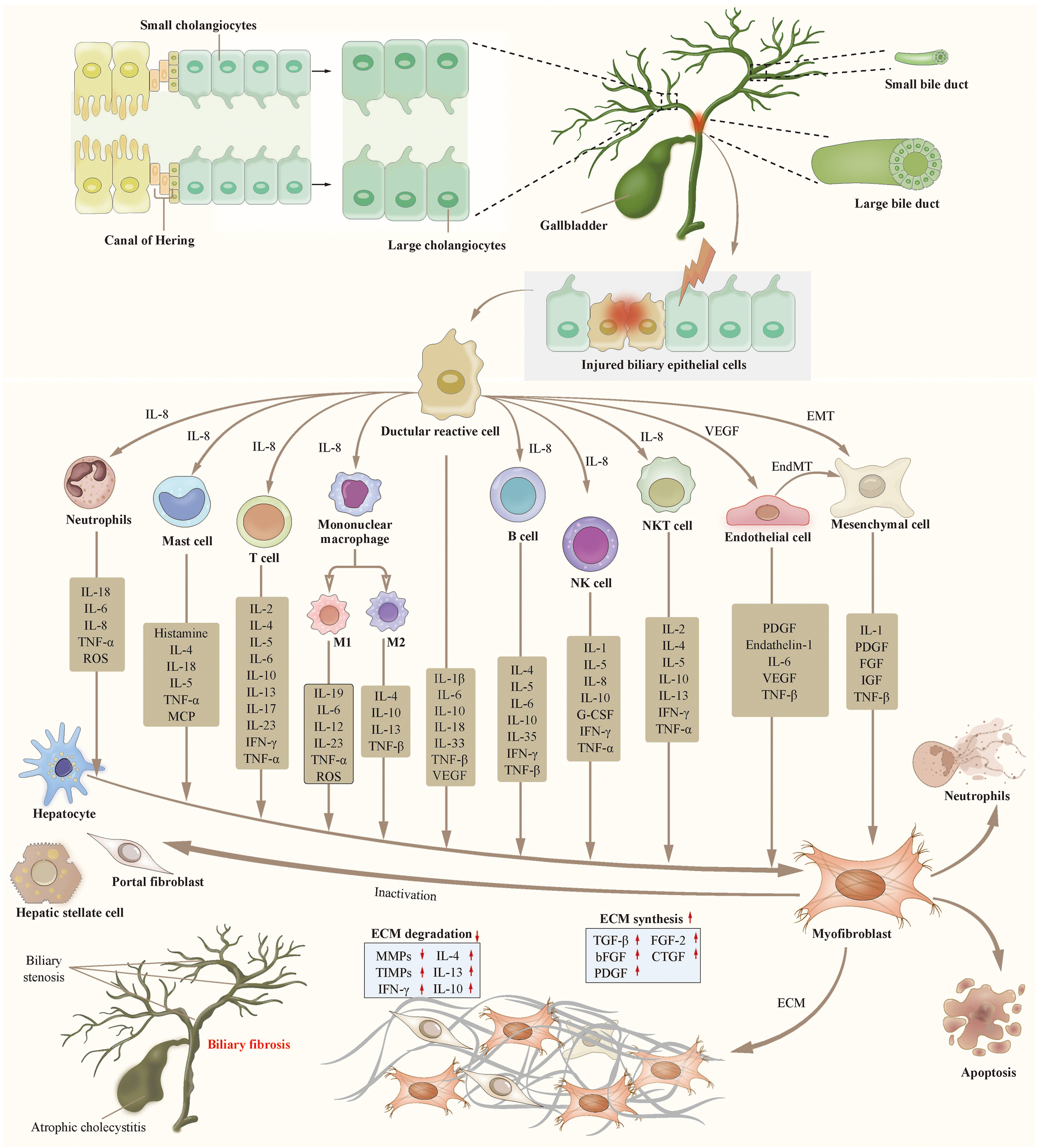
Cellular mechanisms of biliary fibrosis. Biliary fibrosis is a complex process involving multiple cells. Biliary epithelial injury and inflammation lead to the emergence of ductular reactive cell (DRC), which is a subtype of abnormally activated cholangiocyte. DRCs promote the activation of fibroblast precursor cells into myofibroblasts through various immune cells and cytokines. DRCs also promote the proliferation and activation of fibroblasts through the EMT or EndMT pathways. The activated myofibroblasts lead to the disordered synthesis and degradation of ECM, resulting in the occurrence and progression of biliary fibrosis. Abbreviation: DRC, ductular reactive cell. VEGF, vascular epithelial growth factor. EMT, epithelial-mesenchymal transition. EndMT, endothelial-mesenchymal transition. NK cell, natural killer cell. NKT cell, natural killer T cell. IL, interleukin. ROS, reactive oxygen species. TNF, tumor necrosis factor. MCP, monocyte chemoattractant protein. IFN, interferon. G-CSF, granulocyte colony-stimulating factor. PDGF, platelet-derived growth factor. IGF, insulin-like growth factor. FGF, fibroblast growth factor. ECM, extracellular matrix. TGF, transforming growth factor. MMP, matrix metalloproteinase. TIMP, tissue inhibitor of metalloproteinase. CTGF, connective tissue growth factor.teaser-image
Cholangiocyte
Cholangiocytes line the entirety of the biliary tract, stretching from the Canals of Hering to the major papilla. During the development of hepatobiliary system, hepatic progenitor cells (HPC) proximal to the portal interstitium differentiate into bile duct-like cells, forming primitive ductal structures, which subsequently evolve into intrahepatic bile ducts [138, 139]. The formation of intrahepatic bile duct is regulated by spatial and temporal gradients of signals from portal endothelial or mesenchymal cells, including Notch [140], Wnt [141], Hedgehog [142], transforming growth factor β (TGF-β) [143], and fibroblast growth factor (FGF) [144]. A primary cell surface Notch ligand, JAG1 impacts cholangiocyte differentiation and canalicular formation [145]. JAG1 or Notch2 mutations are linked to intrahepatic bile duct loss and fibrosis, as seen in Alagille syndrome [146]. The embryonic development of extrahepatic bile ducts (i.e., common bile duct, cystic duct, gallbladder, and hepatic ducts) slightly differs from intrahepatic bile ducts [139], which may be regulated by SOX9, SOX17, HES1, HNF6, HNF-1β, and HHEX [147, 148].
Cholangiocytes in different regions display heterogeneity in differentiation, metabolism, secretion, and absorption properties, which is likely influenced by the varied distribution of vasculature, oxygen partial pressure, metabolites, and substrates along the biliary tree [108, 139]. Large cholangiocytes are columnar and respond to damage through the cAMP-ERK1/2 signal pathway, small cholangiocytes are cubic and respond to damage via the IP3K/Ca2+ signaling pathway [149]. Cholangiocytes differentiate along the biliary tree from smaller to larger ducts, and exhibit strong plasticity during biliary tract injury and repair. Small cholangiocytes acquire the characteristics of large cholangiocytes and repair the large bile ducts. Cholangiocyte organoids derived from various regions of biliary tract exhibit repair effects on the entire biliary tree [150]. In addition, HPCs within the Canals of Hering and PBGs are able to respond to biliary epithelial injury and loss with proliferation, differentiation, and maturation to restore epithelial integrity [127, 150]. However, the biliary system maintains a balanced state of slow renewal, where hepatocytes and cholangiocytes replenish cells lost to apoptosis predominantly through mitosis rather than relying on the differentiation of HPCs [151, 152]. When normal regeneration mechanisms are impaired, HPCs proliferate and differentiate into hepatocytes and cholangiocytes [153]. Hepatocytes and cholangiocytes specially differentiate into each other in zebrafish models with serious liver injury [154, 155] and in mouse models with disrupted Notch signaling [156].
Cholangiocytes perform essential physiological functions: (1) part of synthesis and secretion of bile; (2) regulating the flow, composition and pH of bile via secretory and absorptive capabilities [149, 157], [158], [159], [160]; (3) constructing a barrier against detrimental molecules and microorganisms via IgA and tight junctions of bile duct cells; (4) releasing immune and blood components into biliary lumen via basement membrane; (5) part of liver regeneration [154, 155]. Moreover, certain studies reported potential roles of cholangiocyte as immune cells [161], [162], [163]. Resting cholangiocytes secrete IgA and antimicrobial peptides, contributing to the homeostasis of the biliary microenvironment. Upon activation, cholangiocytes express HLA-II molecules, adhesion molecules intracellular cell adhesion molecule-1 (ICAM-1) and lymphocyte function associated antigen-3. However, cholangiocytes lack CD80 and CD86 to interact with CD28 and CTLA-4 of T cells, which suggests that HLA-II molecule expression in activated cholangiocytes may be superficial. Research has found that cholangiocyte presented antigens to non-traditional T cells like mucosal-associated invariant T (MAIT) cells and natural killer T (NKT) cells [164], leading to the release of inflammatory cytokines in gut and liver [165].
When biliary tract is injured, resting cholangiocytes are activated to acquire stronger pro-proliferative, pro-inflammatory, and pro-fibrotic abilities, contributing to the recruitment and activation of immune cells, vascular cells, and mesenchymal cells to form a biliary repair complex (also known as ductular reaction). Desmet defined the activated cholangiocytes in the ductular reaction, termed ductular reactive cells (DRC), as “pacemakers of biliary fibrosis” [166]. Contrasting with the biological traits of HPCs and resting cholangiocytes, DRCs exhibit certain morphological and functional mesenchymal cell features. In chronic biliary lesions, DRCs express epithelial mesenchymal transition (EMT) markers like S100A4, vimentin, snail and MMP-2, while downregulating epithelial markers like E-cadherin [167], which is essential for biliary injury repair [168]. Capacity of DRC to repair and reconstruct biliary structures is linked to Notch and Yes-associated proteins (YAP) [169], [170], [171], [172]. Furthermore, studies demonstrated a significant correlation between the abundance of DRCs and portal fibrosis [169, 173]. DRCs are the result of adaptive mechanisms of biliary injury, accumulating with ongoing pathological repair [174]. Interestingly, secretory traits of RDC resemble the senescent-associated secretory phenotype (SASP) of aging cells [175], which suggests that RDCs might promote biliary inflammation and fibrosis via the SASP mechanism [176].
Endothelial cell
Endothelial cells are fundamental components of blood vessels. Injuries to these cells may disrupt substance exchange between blood and tissues, leading to ischemic and metabolic disorders. Proliferation of fibroblast and myofibroblast in fibrotic tissues requires increased nutrients, potentially triggering abnormal angiogenesis. Endothelial cells also contribute to the fibrotic process. Research indicates that endothelial cells from different fibrotic tissues exhibit dissimilarities. In cirrhotic liver tissue, two new endothelial cell subtypes have been identified: plasmalemma vesicle-associated protein + (PLVAP+) and atypical chemokine receptor 1 + (ACKR1+) cells, both promoting leukocyte migration. Endothelial cell subtypes in human biliary system remain undefined. Coagulation-mediated platelet activation inhibited chronic cholestatic liver necrosis and biliary fibrosis through thrombin-mediated protease-activated receptor 4 (PAR-4) in mice [177], which implies that endothelial injury may influence biliary fibrosis progression via platelet activation. Evidence from kidney and heart confirmed endothelial mesenchymal transition (EndMT) contributed to myofibroblast formation [8, 177].
Immune cells
Immune and inflammatory reactions are considered early events of fibrosis. Activated immune cells, including macrophages (Kupffer cells), mast cells, T cells, B cells, NK cells, dendritic cells, and granulocytes, produce cytokines and inflammatory mediators to activate fibroblasts, promoting biliary fibrosis.
Macrophage
Macrophages in the hepatic sinusoids, also known as Kupffer cells, efficiently remove foreign objects, pathogens, and apoptotic or necrotic cells. Activated Kupffer cells secreting a range of cytokines and chemokines, are crucial in the immune response, inflammation, and fibrosis of cholestatic liver diseases [178]. In PBC and PSC, Kupffer cells may undergo classic M1 activation or alternative M2 activation. In the M1 activation pathway, M0 Kupffer cells, stimulated by pathogens or toxins like LPS, differentiate into the M1 subgroup, releasing TNF-α, IL-6, IL-1β, and TGF-β, which promote inflammation, immune response, and biliary fibrosis [179, 180]. Concurrently, TNF-α, IL-6, and IL-1β further stimulate M1 Kupffer cells through autocrine and paracrine pathways to exacerbate inflammation, with IL-6 also triggering hepatocyte and cholangiocyte proliferation by activating PI3K, JAK/STAT, and p38MAPK pathways [181]. In alternative pathways, M2 Kupffer cells, activated by IL-4, IL-10, and IL-13, release IL-4, IL-10, IL-13, and TGF-β, playing roles in anti-inflammation, damage repair response, and fibrosis [179, 180]. Notably, studies have confirmed that the activation of M1 and M2 subgroups of Kupffer cells was regulated by miRNA. miRNA-124 simultaneously inhibits the activation of both M1 and M2 subgroups [182], while miRNA-233 [183] and miNA-155 [184] inhibit the activation of Kupffer cells in M1 and M2 subgroups, respectively. In addition, M2 Kupffer cells induce apoptosis of M1 Kupffer cells through IL-10 pathway to maintain the balance of M1/M2 Kupffer cells, which is crucial for the pathology of cholestatic liver diseases. For instance, one study observed the therapeutic effect of different exogenous bone marrow-derived macrophages (i.e., M0, M1, or M2 macrophages) in cholestatic mice, and found that M0 and M1 macrophages significantly improved liver fibrosis, whereas M2 macrophages showed no therapeutic effect [185]. Another research in BALB/c and C57BL/6 mice showed that M2 Kupffer cells induced M1 Kupffer cell apoptosis and regulated M1/M2 balance by secreting IL-10, thereby protecting against alcohol or high-fat diet induced fatty liver and hepatocyte apoptosis [186]. These evidences suggest that the activation of M2 Kupffer cells and the regulation of M1/M2 balance may be potential targets for the treatment of biliary fibrosis.
Once biliary injury occurs, besides resident Kupffer cells (ResKC), monocyte-derived Kuffer cells (MoKC) are also recruited and activated, however, these two macrophage types display distinct effects. De Muynck et al. investigated the roles of ResKCs and MoKCs in PSC mouse models, and found that CLEC4F+TIM4+ResKCs were depleted after chronic cholestatic liver injury, infiltrating CLEC4F+TIM4- MoKCs were recruited in large quantities during the acute phase of liver injury, and MoKCs expressed higher levels of proinflammatory and proliferative markers than ResKCs [187]. However, further research in experiments with Clec4fDTR transgenic mice indicated that conditional depletion of Kuffer cells before and during early cholestasis induction had no effects on both the hepatic myeloid cell pool composition following injury progression and disease outcomes [187]. In addition, inducing hepatic macrophage depletion seems to benefit the treatment of cholestatic diseases. In a rat model of Glisson intrathecal biliary fibrosis induced by α-nephthyl isothiocyanate (ANIT), liver macrophage depletion alleviated biliary fibrosis by reducing inflammatory mediators, such as MCP-1, IFN-γ, IL-10, and TGF-β1 [188]. Notably, the cholangiocyte-derived lncRNA-H19 plays a key role in macrophage activation, polarization and chemotaxis, through the CCL-2/CCR-2 signaling pathway [189]. LncRNA-H19 may selectively deplete macrophages to inhibit cholangiocyte proliferation and biliary fibrosis in BA patients [190].
Mast cell
Mast cells (MC) activate other innate immune cells such as neutrophils, macrophages/Kupffer cells, dendritic cells, NK cells, and innate lymphocytes by releasing histamine. Histamine facilitates antigen uptake and presentation to adaptive immune cells and promotes cell proliferation, vascular cell activation and fibrosis. Studies in both animal models and human cholestatic diseases indicated significant MC proliferation and increased histamine levels. Research has shown that H1 histamine receptor (H1HR) binding with Gαi triggers a Ca2+-mediated intracellular signal transduction, while H2 histamine receptor (H2HR) binding with Gαs activates the cAMP/ERK signaling pathway. Large cholangiocytes and adjacent MCs express histamine receptors (HRs), inducing paracrine cascades that contribute to disease progression. When the biliary tract is injured, MCs selectively target large cholangiocytes via H2HR-mediated cAMP/ERK1/2 signaling [191]. However, the specific effects of MCs on cholangiocytes remain unconfirmed. Additionally, the elevated SASP expression of cholangiocyte induces MC migration. Notably, MC knockout can reduce liver and bile duct damage in bile duct ligation (BDL) mouse models, highlighting the roles of MC in biliary fibrosis [192].
NK cell
NK cells are key components of innate immunity, which directly attack various target cells, including cancer cells, virus-infected cells, larger pathogens, and transplanted organs and tissues. NK cell cytotoxicity primarily involves four pathways: (1) direct cytotoxic effect through extracellular perforin and granzyme; (2) inducing cell apoptosis through Fas (CD95) ligands and TRAIL molecules; (3) direct and indirect killing effects by releasing cytokines such as IFN-γ, TNF-α, IL1, IL5, IL8, IL10, and granulocyte colony-stimulating factor; (4) antibody-dependent cell-mediated cytotoxicity (ADCC) mediated by CD16 molecules. In addition, NK cells also regulate the activation or inhibition of T-cell responses. Current research suggests that NK cells play dual roles in the development and progression of liver fibrosis, including profibrotic and anti-fibrotic functions [193]. NK cells destroy activated hepatic stellate cells (HSC) and virus infected hepatocytes, thereby ameliorating fibrosis [136]. Regulating NK cells can suppress the activation of HSCs and improve their cytotoxicity against activated HSCs to reduce fibrosis [194].
NKT cell
NKT cells, a subgroup of lymphocytes expressing both the T cell receptor (TCR) and NK cell associated markers, recognize glycolipid antigens presented by the non-polymorphic MHC class I-like protein CD1 [195]. The distribution proportion of NKT cells is up to 10 % of T cells in the liver and less than 1 % in extrahepatic sites [195]. NKT cells release Th1 cytokines (e.g., IFN-γ, IL-2, and TNF-α) and Th2 cytokines (e.g., IL-4, IL-5, IL-10 and IL-13) to polarize immunological responses towards either Th1 or Th2 phenotypes [196]. An imbalance of pro-inflammatory and anti-inflammatory immune responses within the liver is thought to be a major cause of autoimmune liver disease [197], such as AIH, PBC and PSC. NKT cells play multiple roles in autoimmune liver diseases by mediating anti-inflammatory and pro-inflammatory immune responses, or regulating other types of immunoregulatory cells [195]. Aso-Ishimoto et al. [198] investigated the status of NKT cells in the liver and peripheral blood samples of PBC patients with different stages, and found that the proportions of NKT cells were significantly decreased in the liver of patients with early PBC, however, the proportion of CD56+ NKT cells increased in patients with advanced PBC [198]. These findings suggest that activated NKT cells may contribute to cholangiocyte death resulting in the progression of PBC. Research results from Berntsen et al. [199] and Jia et al. [200] also highlighted the role of NKT cells in biliary inflammation and fibrosis.
T cell
Research has confirmed that various T cell subgroups (e.g., Th, Tc and Treg cells) contribute to autoimmune cholangiopathies like PBC, PSC, IgG4-SC, and BA. Chronic nonsuppurative destructive cholangitis (CNSDC) as pathological features of PBC, is the results of small bile duct damage caused by inflammatory cells in the liver. Studies showed that CD8+ and CD4+ T cells are predominant in portal tract inflammation of PBC [70], with cytokine environments varying across disease stages [201]. The Th1 cell environment of early stages gradually shifts to Th17 cells of advanced stages. A decline in Treg cells infiltrating the portal area and an imbalance in cytotoxic T cell proportions correlate with PBC progression. Clinical studies indicated a higher proportion of total Treg cells in hepatic hilum lymph nodes of BA patients than in peripheral blood, and proportions of activated Treg and CD4+ T cells in these lymph nodes are significantly elevated [202]. Mechanistic studies revealed that biliary destruction of BA was predominantly caused by CD4+ and CD8+ T cells [203]. Interactions between Th1 and Treg cells regulate biliary fibrosis of BA via the IFN-γ/STAT signaling pathway [204].
T cells also play crucial roles in parasitic infection-induced biliary fibrosis. Th1, CD4+ Th2, Th17, and Treg cells are implicated in biliary fibrosis of Clonorchis sinensis-infected mice [205]. Th1 cells inhibit HSC activation by secreting IFN-γ, thus preventing biliary fibrosis. Th2 cytokines such as IL-4, IL-5 and IL-13 promote fibrosis by activating HSCs, while Th17 cells trigger liver granulomas and fibrosis via IL-17 pathway [206]. Furthermore, studies showed that miRNA-214 knockout or downregulation reduced the number of T cell and pro-fibrotic cytokine-mediated perivascular fibrosis in mice [207]. Low-dose IL-2 treatment via Treg cells mitigates biliary injury and fibrosis in sclerosing cholangitis models [208].
B cell
The involvement of B cells in PBC, PSC and IgG4-SC is well-established [209]. The pathogenesis of B cells in immune-related hepatobiliary diseases involves producing autoreactive antibodies, and recruiting T cells via cytokines and antigen-presenting cells (APC) [210]. CD19+ B cells in PBC release higher levels of IL-6 and TNF-α than healthy controls, which suggests that B cells contribute to inflammatory environment in PBC, leading to persistent inflammation and fibrosis progression [210]. Furthermore, serum from PSC patients triggers cholangiocytes to release inflammatory mediators like IL-6 [210]. Histologically, B cells in IgG4-related hepatobiliary diseases resemble those in fibrotic bile ducts, and patient-derived B cells with IgG4 related disease (IgG4-RD) can stimulate human primary pancreatic fibroblasts to produce collagen [211]. Notably, plasma cells, rather than naive or memory B cells, are most effective at producing collagen during coculture [211]. A study of six IgG4-SC patients revealed oligoclonal amplification of IgG4+ clones in peripheral blood mononuclear cells, with these clones predominating in tissue of IgG+ cell infiltration [212]. This indicates a direct pathogenic role for oligoclonal amplified B cells in IgG4-SC. In addition, B cell depletion therapy is recognized as an effective treatment for IgG4-RD [212], which reduces disease activity, fibrosis markers, ELF score, and the number of myofibroblasts [53].
Fibroblast and myofibroblast
Fibroblast and myofibroblast are the same cells in different functional states. Normally, fibroblasts form connective tissue by synthesizing ECM components (i.e., collagen, proteoglycan, elastin, fibronectin, microfibrils and laminin), and reshaping the microstructure of ECM through covalent cross-linking, protein glycosylation and balanced secretion of matrix metalloproteinases (MMPs) and matrix metalloproteinase inhibitors (MMPIs) [213]. Fibroblasts exhibit heterogeneity across tissues and organs, and maintain stable lineages within the same tissue. Single-cell RNA sequencing data revealed less than 20 % of gene overlapped among fibroblasts from mouse hearts, skeletal muscles, intestines, and bladders [214]. Specific ECM components and fibroblast micro-remodeling activities create microenvironments for tissue-specific cells such as keratinocytes in skin, epithelial cells in lung, striated muscle fibres in skeletal muscle, and endothelial cells in blood vessels [215]. Fibroblasts exert a “dragging” effect on the surrounding ECM, which is a unique biomechanical regulatory mechanism promoting tissue-level mechanical forces and matrix polarization [216]. Additionally, fibroblasts serve as precursor cells for specific mesenchymal cells such as osteoblasts and adipocytes, and can divide into fibroblasts and other tissue-specific mesenchymal cells [217, 218]. A cross-tissue mouse study revealed two common fibroblast subgroups expressing Pi16 or Col5a1, and further tracing of the genetic lineage revealed that the latter is more prevalent in PDGFR-α+ fibroblasts [219]. This suggests that fibroblasts persist through injury, tumorigenesis, or inflammation, forming specialized subgroups. Notably, cell subgroups similar to Pi16+ mouse fibroblasts have been observed in human tissues as well [219], indicating cross-species similarity in fibroblast lineages.
Myofibroblast activation and excessive ECM accumulation are key events in fibrosis. Normally, myofibroblasts are few in number. In response to tissue damage or structural changes, myofibroblast precursor cells transform into myofibroblasts. Under physiological conditions, myofibroblasts express α-SMA and other contractile proteins, coordinating biomechanical remodeling and contraction through traction on many newly secreted ECMs, thereby achieving physiological repair and remodeling of injured tissue [220], [221], [222]. In addition, myofibroblasts also alter the surrounding tissue environment by regulating the function of resident immune cells [220, 223, 224] and phagocytosing necrotic cells [225]. However, when tissues are severely and repeatedly damaged or continuously stimulated by inflammation, abnormal activation of myofibroblasts leads to excessive ECM synthesis and accumulation, promoting scar formation and fibrosis, resulting in organ dysfunction, and even the development of tumour [4].
The origin of myofibroblasts remains controversial. Fibroblast residents in the tissue are considered the primary source of myofibroblasts after injury, and other specialized mesenchymal cells can also change their lineage specificity and generate myofibroblasts. For instance, some myofibroblasts in injured skin or other tissues originate from circulating bone marrow hematopoietic progenitor cells [226], [227], [228]. Mice experiments found that mature adipocytes in the skin, myoblasts in skeletal muscle, and mesenchymal cells around blood vessels can be transformed into fibroblasts [137, 229], [230], [231], [232], [233]. Even, the endothelial and epithelial cells of the heart have been confirmed to transform into neighbouring myofibroblasts in cardiac fibrosis through the EMT and EndMT pathways [13]. In biliary fibrosis, potential myofibroblast precursors include liver stem cells [234], hepatocytes [235, 236], cholangiocytes [237, 238], portal fibroblasts [239], [240], [241], HSCs [242], perivascular cells [243], and bone marrow-derived cells [244]. Inhibiting the transformation of these precursor cells into myofibroblasts is crucial for antifibrotic therapy.
The activation of myofibroblasts is complexly regulated by various fibrosis regulatory factors such as TGF-β [176, 237, 245], [246], [247], [248], [249], [250], [251], [252], [253], FGFs [254], [255], [256], platelet derived growth factors (PDGFs) [176, 257], [258], [259], connective tissue growth factor (CTGF) [260, 261], peroxisome proliferators-activated receptor (PPAR) [262], [263], [264], Wnt [265], non-phagocytic cell oxidase (NOX) [266], endoplasmic reticulum (ER) [267], a-kinase anchor protein 12 (AKAP12) [268], cellular communication network factor (CCN) [269, 270], Mesothelin [239], and ubiquitin c-terminal hydrolase L1 (UCHL1) [271]. Single-cell transcriptome sequencing revealed that the fibroblast to myofibroblast transformation involved transcriptional changes akin to cell differentiation, indicating that they also follow a hierarchical lineage structure [226, 272], [273], [274]. However, this lineage does not follow the typical stem cell lineage pattern, where undifferentiated and long-term self-renewal progenitor cells unidirectionally differentiate specific tissue cells [275]. Increasing evidence suggests that myofibroblasts are not terminally differentiated, with their transformation from fibroblasts being temporary and reversible under physiological conditions [276]. When the tension of wounds decreases, the expression and function of contraction genes in myofibroblasts will decrease. Once the injury heals, activated myofibroblasts transform into fibroblasts or gradually decrease through apoptosis and senescence pathways [277, 278]. An animal study found that myofibroblasts in mouse skin wounds can transform into lipid-filled adipocytes [279]. Notably, mature myofibroblasts adhere to the ECM longer than fibroblasts under the same signals [280], suggesting that detachment is crucial for terminating the myofibroblast state. Tension-sensitive myocardin-related transcription factor A (MRTF-A) and serum reactive factor (SRF), activated by adhesion signals, may facilitate this process.
Biliary and intestinal microbiota
Microorganisms play crucial roles in development and progression of disease. Numerous studies have identified specific microbial changes in cholangiopathies such as PSC [281], [282], [283], [284], [285], [286], [287], [288], [289], [290], [291], [292], [293], [294], [295], [296], PBC [297], [298], [299], [300], [301], [302], [303], BA [304, 305], IgG4-SC [292] and SSC-CIP [306] (Table 4). Particularly, Sabino et al. reported a notable gut flora characteristic of PSC patients was reduced bacterial diversity with a significantly higher proportion of Enterococcus, Clostridium, and Lactobacillus [281]. Other studies discovered increased abundance of Proteobacteria, Bacteroidetes, and Paramonas in feces of PSC patients [307, 308]. Ehud Zigmund et al. reported that bile bacterial culture of 189 PSC patients was positive and biliary Enterococci colonization was an independent risk factor for PSC progression [80].
Characteristics of bile and gut microorganisms in biliary fibrosis-related cholangiopathies.
| Cholangiopathy | Sample | Changes of microbiome | References | |
|---|---|---|---|---|
| Increased | Decreased | |||
| PSC | Feces |
Enterococcus genus
Fusobacterium genus Lactobacillus genus Veillonella genus Rothia genus Streptococcus genus Parabacteroides genus Ruminococcus genus Proteobacteria genus Bacteroides genus Exophiala genus Enterobacteriaceae Clostridium genus |
Adlercreutzia equolifaciens genus
Prevotella copri genus Saccharomyces cerevisiae genus Firmicutes (except Veillonella genus) Eubacterium Ruminococcus Bifidobacterium Lactobacillus Coprococcus Faecalibacterium Phascolarctobacterium Bacteroides thetaiotaomicron Faecalibacterium prausnitzii |
Sabino et al. [281] Kummen et al. [282] Bajer et al. [283] Iwasawa et al. [284] Rühlemann et al. [285] Torres et al. [286] Rühlemann et al. [287] Vieira-Silva et al. [288] Lemoinne et al. [289] Kummen et al. [290] Denoth L et al. [291] Liu Q et al. [292] Lapidot Y et al. [293] Cortez et al. [294] |
| Bile |
Fusobacteria
Enterococcus Proteobacteria Staphylococcus Neisseria |
Unclear | Miyabe K et al. [295] Liwinski et al. [296] |
|
| PBC | Feces |
Bifidobacterium
Streptococcus Lactobacillus Enterococcus Serratia Klebsiella Veillonella Clostridium Pseudomonas Haemophilus Weisella Enterobacter asburiae Haemophilus parainfluenzae Megasphaera micronuciformis Paraprevotella clara |
Clostridiales
Faecalibacterium Ruminococcaceae Oscillospira Sutterella Bacteroides Lachnobacterium Bacteroides eggerthii Ruminococcus bromii |
Furukawa M et al. [297] Yijun Zhou et al. [298] Tang et al. [299] Chen et al. [300] Lammert et al. [301] Abe et al. [302] Lv et al. [303] |
| BA | Feces |
Klebsiella
Streptococcus Veillonella Enterococcus Proteobacteria |
Bacteroidetes
Bifidobacterium |
Song W et al. [304] Wang J F et al. [305] |
| IgG4-SC | Feces | Unclear | Blautia | Liu Q et al. [292] |
| SC-CIP | Feces |
Rothia dentocariosa
Enterococcus faecium |
Unclear | Blesl a et al. [306] |
-
PSC, primary sclerosing cholangitis; PBC, primary biliary cholangitis; BA, biliary atresia; IgG4-SC, IgG4-related sclerosing cholangitis; SC-CIP, sclerosing cholangitis in critically ill patient.
The causal relationship between biliary fibrosis and microbiota remains complex. It has been confirmed that the microbiota contributed to the occurrence and progression of biliary fibrosis through multiple pathways such as inducing mucosal inflammation and regulating host immunity and metabolism. Biliary epithelial damage and chronic mucosal inflammation caused by biliary flora colonization initiate the biliary fibrosis process [80]. LPS of bacteria induces TGF-β1 expression and the formation of bipolar cells with fibroblast morphology in intrahepatic bile ducts [309]. H. pylori in the biliary tract stimulates reactive oxygen species (ROS) production, fostering cholangiocyte proliferation and biliary fibrosis [79]. The gut microbiota facilitates bile acid metabolism by increasing fecal bile acid excretion, stimulating intestinal GLP-1 secretion, and alleviating hepatobiliary injury in Mdr2−/− mice [310]. Notably, microbiota is associated with certain genetic risk sites of PSC, such as fucosyltransferase 2 (FUT2) [311]. In addition, research examining fecal metabolic characteristics in peribiliary fibrosis patients with liver flukes showed significantly increased concentrations of ethanol, glycine, tyrosine, and N-acetylglucosamine, suggesting crosstalk between host and gut microbiota cometabolism in liver fluke-induced biliary fibrosis [312]. However, research on the impact of biliary fibrosis on intestinal microbiota is limited. Biliary fibrosis may alter the characteristics of the gut microbiota through bile acid metabolism. There is a lack of longitudinal studies demonstrating the dynamic changes in microbiota during biliary fibrosis.
Signaling pathways related to biliary fibrosis
Biliary fibrosis involves essential signaling pathways, including developmental-related signaling pathways, growth factor-related signaling pathways, bile acid-related signaling pathways, and immune and inflammatory signaling pathways [313]. We describe transduction process and biological effects of these signals in biliary fibrosis in Figure 3.
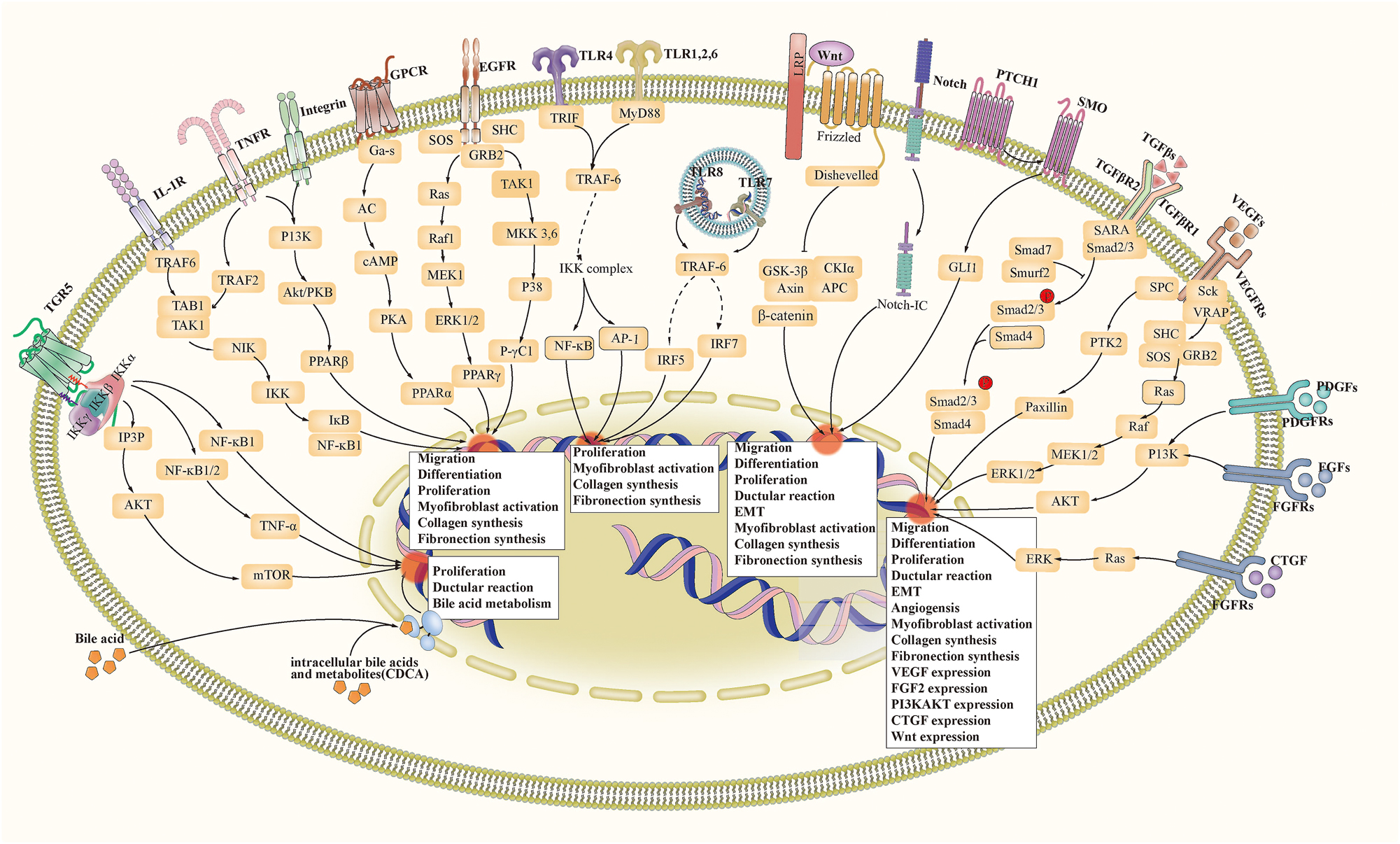
Signaling pathways related to biliary fibrosis. Abbreviation: ECM, extracellular matrix. EMT, epithelial-mesenchymal transition. TGR5, Takeda G protein-coupled receptor 5. TNF, tumour necrosis factor. GPCR, G-protein-coupled receptor. EGFR, epidermal growth factor receptor. TLR, toll-like receptor. LRP, low-density lipoprotein receptor-related protein. TGF(R), transforming growth factor (receptor). VEGF(R), vascular endothelial growth factor (receptor). PDGF(R), platelet-derived growth factor (receptor). FGF(R), fibroblast growth factor (receptor). CTGF(R), connective tissue growth factor (receptor). FXR, farnesoid X receptor. PPAR, peroxisome proliferator-activated receptor.
Signaling pathways related to biliary development
The Notch [314], Wnt [315], and Hedgehog [316] pathways are pivotal in embryonic and organ development, crucial for cell proliferation and differentiation, tissue and organ formation, adult stem cell maintenance, and post-injury repair and reconstruction [142, 317], [318], [319]. Abnormal activation of these signaling pathways is closely related to biliary fibrosis in cholestatic liver disease. In detail, abnormal Notch signaling is linked to irregular biliary development and fibrosis [318]. JAG1 mutations in the Notch pathway cause Alagille syndrome [320]. Prolonged Notch signaling can lead to liver fibrosis in a high-fat diet-induced fatty liver mouse model, likely due to abnormal activation of HSCs by activated Notch signaling [321]. Abnormal activation of Wnt signaling pathway promotes fibrosis in various organs [322], [323], [324] by inhibiting proliferation of myofibroblast or inducing myofibroblast apoptosis [323]. Moreover, inhibition of Hedgehog pathway can reduce biliary fibrosis in BDL-induced mice [325].
Growth factor-related signaling pathways
TGF-β signaling pathway
TGF-β exerts multiple biological effects, including regulating cell growth and plasticity, activating myofibroblasts, promoting immune tolerance, and suppressing inflammation. Target cells of TGF-β include epithelial, immune, fibroblast, vascular, and nerve cells [247]. Despite TGF-β affecting various target cells, its overarching goal is to repair injury and maintain tissue integrity [326]. Fibrosis is among the most prevalent disease processes associated with TGF-β [247].
The exact mechanism of TGF-β activation remains unclear, the hypothesis that the integrin family activates TGF-β has gained credibility through multiple verifications [326]. TGF-β, primarily derived from macrophages, epithelial cells, and fibroblasts, plays pivotal roles in the initiation and progression of fibrosis by influencing fibroblasts and epithelial cells [327]. TGF-β regulates fibroblast activation at nearly all stages of tissue damage and repair. The activated TGF-β1/Smad3 signaling pathway recruits fibroblasts to injury sites and triggers their transformation into myofibroblasts. These activated myofibroblasts are crucial for ECM synthesis and remodelling and act as signaling regulators for epithelial, immune, and endothelial cells. TGF-β stimulates the production of enzymes and molecular chaperones related to fibrillar collagen, facilitating cross-linking and inhibiting degradation of ECM. TGF-β also enhances ECM synthesis and collagen cross-linking through feedforward regulation. Furthermore, activated TGF-β fosters self-expression in both autocrine and paracrine pathways, and stimulates more TGF-β activation by inducing integrin expression on fibroblasts and epithelial cells. TGF-β is also a dominant regulator of EMT and cell senescence and apoptosis [328].
PDGF signaling pathway
Under normal conditions, PDGF is predominantly expressed in platelet α granules and facilitates cell division and proliferation. During biliary injury, PDGF is synthesized and released by platelets, vascular endothelial cells, pericytes, and Kupffer cells, influencing the proliferation of fibroblasts and vascular endothelial cells [329]. The combination of PDGF and PDGF receptor (PDGFR) activates downstream signaling pathways like RAS/MAPK, PI3K/AKT, JAK/STAT, PLC-γ, and Rho, to promote collagen synthesis and cell adhesion [329]. An animal study indicated that chronic bacterial colonization in rat bile ducts induced biliary fibrosis through a PDGF-dependent pathway [330].
FGF signaling pathway
The fibroblast growth factor (FGF) family comprises of at least 20 members, from FGF-1 to FGF-20, divided into seven subfamilies [256]. Each subfamily functions through distinct pathways: paracrine (FGF-1, FGF-4, FGF-7, FGF-8, FGF-9), endocrine (FGF-19), and intracellular pathway (FGF-11) [256]. FGF binding to FGF receptor initiates FGF signal transduction. In paracrine pathways, FGF activation involves heparin and heparan sulfate as cofactors. In endocrine pathways, α Klotho or β Klotho aid FGF activation of FGFR. Endogenous FGFs exert direct effects independent of receptors. The FGF signaling pathway is pivotal in embryonic development, angiogenesis, wound healing, tissue repair, and organ regeneration [331]. The endocrine FGF19 subfamily also regulates bile acid and cholesterol balance via a Klotho protein-dependent pathway. However, the exact roles of the FGF family in biliary fibrosis remain to be fully elucidated.
VEGF signaling pathway
VEGF is a highly active mitogen for endothelial cells [332], targeting endothelial cells to stimulate angiogenesis, increase vascular permeability, and prevent endothelial cell apoptosis [333]. Research indicated that the VEGF family played roles in pathological processes like liver fibrosis, including the regulation of angiogenesis during tissue collagen deposition and balancing nutrient supply [334].
CTGF signaling pathway
CTGF is a newly recognized growth factor related to PDGF, which enhances fibroblast proliferation and chemotaxis, and regulates cell migration and differentiation [335]. Numerous studies have identified that CTGF was associated with organ fibrosis, including blood vessels, skin, heart, kidney, pancreas, lung, and liver [335]. CTGF is a key profibrotic cytokine in liver fibrosis, which directly promotes the proliferation, migration and activation of HSC, resulting in excessive synthesis and accumulation of ECM (e.g., type I and type III collagens) [336]. Furthermore, CTGF interacts with other profibrotic factors to amplify their functions. For instance, the binding of CTGF and TGF-β1 is a critical step for TGF-β1 to manifest profibrotic effects [335].
Downstream signaling pathways of growth factors
The PI3K/AKT signaling pathway connects to upstream receptors such as RTKs, Toll-like receptors (TLR), B cell receptors (BCR), GPCRs, and JAK/STAT, and targets downstream molecules such as mTOR, GSK3, FOXOs, TSC2, and MDM2, to regulate cell cycle, metabolism, angiogenesis, and inflammation.
The JAK/STAT signaling pathway facilitates signal transduction within the nucleus [337]. JAK/STAT signaling is influenced by the activation of the PI3K/AKT/mTOR pathway and PDGFs. Studies have shown that JAK2/STAT3 signaling played a role in PDGF-induced fibrosis. Additionally, the activation of JAK/STAT signaling is vital for the production of TGF-β [338].
The Wnt/β-catenin pathway activates and interacts with TGF-β1, mediating myofibroblast activation and inducing the expression of vimentin, type I collagen, and fibronectin [339].
Immune and inflammatory signaling pathways
TLR signaling pathway
TLRs recognize pathogen-associated molecular patterns (PAMP), which are highly conserved components of microorganism [340]. These receptors detect various microbial stimuli, recruit specific adaptor proteins, initiate signaling cascades, and trigger immune responses against pathogens, linking innate and adaptive immunity. TLR ligands include synthetic agonists and microbial components like LPS, peptidoglycan (pGN), lipoteichoic acid (LTA), and lipoarabinomannan (LAM). Specific TLRs recognize corresponding ligands. For instance, TLR2 identifies lipoproteins and peptidopolysaccharides, including pGN and LTA, while TLR4 recognizes LPS [340].
Animal studies have highlighted the role of TLR signaling in biliary fibrosis, especially in microbial and parasitic-related cholangiopathies. TLR2 activates TGF-β1-Smad2/3 through the AKT-p38 pathways, leading to IL-6 secretion and promoting myofibroblast activation and biliary fibrosis in mice infected with C. sinensis [76, 341, 342]. TLR9 stimulates cholangiocytes to secrete IL-6 and TNF-αvia the ERK pathway, exacerbating biliary inflammation and fibrosis [343]. TLR9 deficiency reduces MCP-1 and collagen expression, and mitigates biliary fibrosis in BDL mice [344]. Furthermore, Yan et al. found that TLR4 deficiency worsened biliary injury and fibrosis of mice caused by C. sinensis, possibly due to an enhanced type II immune response and proinflammatory effects [77].
PPAR signaling pathway
PPARs comprise of three homologous variants: PPAR-α, PPAR-β/δ, and PPAR-γ. Upon activation by various natural or synthetic ligands, PPARs bind to the retinoid X receptor (RXR) to form a heterodimer. This formation facilitates the recruitment of coactivators and the release of corepressors, thereby regulating the expression of genes involved in lipid and glucose metabolism [345]. Notably, PPARs also inhibit the expression of certain genes without directly binding to DNA, which might explain their role in inhibiting inflammation and fibrosis. Activation of PPAR-γ impedes inflammatory responses by inhibiting NF-κB signaling and diminishing the expression of TNF-α and IL-1β in monocytes and macrophages. Dual activation of PPAR-γ/α effectively ameliorates nonalcoholic steatohepatitis by reducing inflammation, steatosis, and fibrosis [346]. The agonists of PPAR-α and PPAR-γ have demonstrated effectiveness in animal models of cardiac [347], renal [348], and pulmonary [349] fibrosis.
Bile acid-related signaling pathways
Enterohepatic circulation of bile acid
Bile acids are synthesized in the liver via two pathways: the classical pathway, predominantly in hepatocytes catalyzed by cholesterol 7α-hydroxylase (CYP7A1), and the alternative pathway, mainly in cholangiocytes and macrophages involving cytochrome P450 family 27 subfamily A member 1 (CYP27A1) and oxysterol 7α-hydroxylase (CYP7B1) [350]. Bile acids are transported to the bile duct through the hepatocyte surface bile acid transport proteins, with some secretion via the multi-drug-resistant protein 2 (MRP2). After gallbladder storage, bile acids enter the intestine, where approximately 95 % are reabsorbed through the apical sodium-dependent bile acid transporter (ASBT) and organic solute transporter α/β (OST-α/β), and then recirculated to hepatocytes via Na+-taurocholate cotransporting polypeptides (NTCP) and organic anion-transporting polypeptides (OATP) [351]. Among them, NTCP is responsible for the uptake of almost all conjugated bile acids and is regulated by farnesoid X receptor – small heterodimer partner (FXR/SHP), while OATP is mainly responsible for the uptake of unconjugated bile acids and is regulated by FXR and PXR. Any obstacle in the enterohepatic circulation of bile acids, such as intrahepatic bile duct loss, stenosis or obstruction, or bile acid transporter dysfunction, may lead to cholestatic liver injury and promote the occurrence and progression of biliary fibrosis. Conversely, biliary fibrosis further affects bile acid metabolism, forming a vicious cycle.
FXR signaling pathway
FXR is a nuclear receptor, expressed in the liver and intestines, where it regulates bile acid and lipid-glucose metabolism. Activated by specific bile acid metabolites such as chenodeoxycholic acid (CDCA), cholic acid (CA), deoxycholic acid (DCA), and lithocholic acid (LCA) [352], FXR plays a regulatory role in bile acid metabolism. Elevated bile acid levels in hepatocytes trigger FXR activation, which then modulates the expression of FXR target genes. This results in reduced bile acid synthesis and reuptake, and increased extracellular bile acid transport [353]. Inhibition of FXR expression inhibits bile acid-induced EMT of cholangiocytes. However, FXR has shown protective effects in renal and liver fibrosis. In animal models of kidney disease, FXR activation improves triglyceride accumulation and proteinuria and reduces ECM accumulation [354]. In fatty liver disease, FXR activation attenuates liver injury by inhibiting the activation of NACHT, LRR, and NLRP3 [355]. In addition, FXR directly regulates the expression of FGF19, which regulates glucose-lipid metabolism and inhibits the progression of liver fibrosis [356].
TGR5 signaling pathway
TGR5, also known as GPBAR1, is mainly expressed in the primary cilia of cholangiocytes [357], and is activated by secondary bile acids and nonconjugated bile acids (LCA>DCA>CDCA>CA) [358]. Low cAMP level and extracellular ERK activation in ciliated cholangiocytes inhibit cholangiocyte proliferation. Conversely, high level of cAMP in non-ciliated cholangiocytes promotes cholangiocyte proliferation [157]. These findings indicate that structurally or functionally deficient cilia may activate the TGR5 and increase intracellular cAMP, leading to various cholangiopathies characterized by excessive proliferation of cholangiocyte, such as BA. In BDL mouse models, activation of the EGFR and ERK pathways mediated by TGR5 promotes cholangiocyte proliferation, and TGR5 knockout reduces cholangiocyte proliferation [359], suggesting that bile acids activate TGR5 to promote the proliferation of cholangiocytes, leading to ductular reactions and ultimately progressing to biliary fibrosis.
However, some studies reported the protective role of TGR5 in cholangiopathies. The expression level of TGR5 is downregulated in PBC and PSC patients [360]. Hov et al. found five nonsynonymous mutations (i.e., W83R, V178M, A217P, S272G, and Q296X) in the TGR5 gene sequencing data of 276 PSC patients and 274 healthy individuals [361]. In addition, TGR5 agonists positively promote bile flow and inhibit cholangiocyte apoptosis [362]. In cholangiopathy mouse models, dual agonists of FXR and TGR5 can improve biliary fibrosis, but selective TGR5 or FXR agonists cannot improve biliary fibrosis [363]. Notably, the opposite effects of TGR5 in different cell subgroups may be related to the release of pro-inflammatory and anti-inflammatory cytokines, meaning that differences in the inflammatory microenvironment have a specific impact on the function of TGR5 [364].
Epigenetic effects
Epigenetics is a reversible and heritable process that involves changes in gene expression patterns caused by mechanisms other than DNA sequence changes. These mechanisms include DNA modification (e.g., methylation), noncoding RNA (ncRNA) mediated gene regulation (e.g., miRNA, lncRNA, and siRNA), histone modification, and chromatin remodelling. These mechanisms can independently or synergistically influence the expression of fibrosis-related genes.
The epigenetic effects are closely related to the occurrence and progression of fibrosis. During non-scar kidney repair, RASAL1 (a promoter encoding Ras tumour protein inhibitor) undergoes hypermethylation, leading to irreversible transcriptional inhibition, persistent myofibroblast activation, and chronic fibrosis. In tissue injury repair, some miRNAs affect the activation, maintenance and termination of myofibroblast, thereby participating in the fibrosis process. LncRNA competitively binds to fibrosis-related miRNAs, regulating the synthesis of ECM. Several specific miRNAs [251, 263, 325, 365], [366], [367], [368], [369], [370], [371], [372], [373], [374], [375], [376], [377], [378], [379], [380], [381], [382] and lncRNAs [189, 190, 383], [384], [385] play essential roles in biliary fibrosis (Table 5). Serum miRNA sequencing analysis of BA patients found 146 differentially expressed miRNAs, with 15 of them (seven upregulated, eight downregulated) predicted to be linked to biliary fibrosis, differentiation, and development [386]. Further qPCR analysis on independent sample sets revealed that miRNA-140-3p was significantly downregulated in BA patients and is expected to become a biomarker for BA diagnosis [386]. Yan et al. analyzed the differential expression of miRNAs in mice infected with C. sinensis, and found 143 downregulated miRNAs and 206 upregulated miRNAs, and further analysis showed that these abnormal miRNAs might regulate signaling pathways related to tumour, TGF-β/Smad7, MAPK, TLR, and PI3K/AKT [387]. Another study found that lncRNA-H19 level was closely related to the severity of liver fibrosis in both PSC/PBC mouse models and patients [388]. LncRNA-H19 deficiency alleviates ductal reaction and biliary fibrosis in BDL mice, and related mechanism might involve S1PR2/SphK2 and let-7/HMGA2 axes [384]. Therefore, the pathogenesis of biliary fibrosis involves various ncRNAs and epigenetic modifications. One ncRNA may simultaneously affect the expression of several mRNAs and regulate multiple cellular events, and the molecular mechanisms of ncRNA in biliary fibrosis have yet to be fully elucidated.
The role of ncRNAs in biliary fibrosis.
| ncRNA | Main effects | Reference |
|---|---|---|
| miRNA-34a | Promoting ductular reaction and cholangiocyte senescence. | [365] |
| miRNA-1295b-3p | Inhibiting expression of ARRB1. | [366] |
| miRNA-148-5p | Inhibiting notch signaling pathway and HSC activation. | [367] |
| miRNA-29a | 1) Regulating PI3KP85α; Inhibiting HSC activation; 2) Regulating ER homeostasis; promoting DNA hypomethylation; 3) Regulating TGF-β and Wnt/β-catenin signaling pathways. |
[368], [369], [370], [371], [372] |
| miRNA-29b1 | Inhibiting HSC activation. | [325] |
| miRNA-98 | Inhibiting HSC activation via HIF-1α/TGF-β/Smad2/3 signaling pathway. | [373] |
| miRNA-21 | 1) Promoting necroinflammation through CDK2AP1; 2) Promoting HSC activation through NLRP3/IL-1β axis. |
[374], [375], [376] |
| miRNA-125b | Promoting cholangiocyte proliferation by upregulating VEGF. | [377] |
| miRNA-let7 | Attenuates ductular reaction by inhibiting NF-κB and IL-13 signaillng pathways. | [378] |
| miRNA-19b | Inhibiting HSC activity and inducing HSC apoptosis. | [379] |
| miANA-150 | Inhibiting HSC activation. | [380] |
| miRNA-155 | Promoting biliary injury by activating JAK2/STAT3. | [381] |
| miRNA-194 | Inhibiting HSC activation. | [380] |
| miRNA-130a/b | Promoting HSC activation | [263] |
| miRNA-340-5p | Inhibiting HSC activation and proliferation. | [382] |
| miRNA-134 | Promoting HSC inactivation. | [251] |
| lncRNA-ACTA2-AS1 | Regulating ductular reaction. | [383] |
| lncRNA-H19 | 1) Inducing activation, polarization and chemotaxis of macrophages; 2) Promoting cholangiocyte proliferation; 3) Promoting HSC differentiation and activation. |
[189, 190, 384, 385] |
-
ncRNA, non-coding RNA; miRNA, microRNA; lncRNA, long non-coding RNA; ARRB1, β-arrestin 1; HSC, hepatic stellate cell; ER, endoplasmic reticulum; TGF-β, transforming growth factor β; VEGF, vascular endothelial growth factor.
Therapeutic strategies for biliary fibrosis-related cholangiopathies
Currently, liver transplantation remains the sole definitive treatment for advanced stages of fibrosis-related cholangiopathies such as PBC, PSC, and IgG4-SC. ERCP provides some assistance in solving the fibrotic stenosis of extrahepatic bile ducts. The drug treatment of biliary fibrosis is mainly etiological and symptomatic treatment like Ursodeoxycholic acid (UDCA) and Obeticholic acid (OCA), lacking specific anti-fibrosis therapy. Based on the latest progress in clinical trials (Table 6) [389], [390], [391], [392], [393], [394], [395], [396], [397], [398], [399], [400], [401], [402], [403], [404], [405], [406], [407], [408], [409], [410] and animal experiments, we summarized the therapeutic strategies for fibrosis-related cholangiopathies.
The interventional clinical trials on the treatment of biliary fibrosis-related cholangiopathies in recent 5 years.
| Cholangiopathy | Interventions | Sample size | Phase | Recruitment status | Year | NCT |
|---|---|---|---|---|---|---|
| BA | Meropenem | 160 | Ⅳ | Completed | 2020 | NCT04370145 |
| Desflurane | 165 | Ⅳ | Not yet recruiting | 2023 | NCT05783518 | |
| Odevixibat | 180 | Ⅲ | Recruiting | 2022 | NCT05426733 | |
| Odevixibat | 245 | Ⅲ | Recruiting | 2020 | NCT04336722 | |
| Norepinephrine bitartrate | 40 | Ⅲ | Recruiting | 2022 | NCT05521152 | |
| OCA | 144 | Ⅱ/Ⅲ | Not yet recruiting | 2023 | NCT06121375 | |
| Maralixibat | 75 | Ⅱ | Completed | 2020 | NCT04524390 | |
| Filgrastim | 400 | Ⅱ | Recruiting | 2020 | NCT04373941 | |
| UC-MSC | 34 | Ⅰ/Ⅱ | Recruiting | 2020 | NCT04522869 | |
| N-acetylcysteine | 12 | Ⅰ | Not yet recruiting | 2024 | NCT06260566 | |
| BMMC | 12 | Ⅰ | Completed | 2022 | NCT05517317 | |
| PSC | Simvastatin | 700 | Ⅲ | Recruiting | 2019 | NCT04133792 |
| Bezafibrate | 104 | Ⅲ | Recruiting | 2020 | NCT04309773 | |
| Ritivixibat | 24 | Ⅱ | Recruiting | 2022 | NCT05642468 | |
| CS0159 | 50 | Ⅱ | Recruiting | 2023 | NCT05896137 | |
| Aramchol meglumine | 24 | Ⅱ | Not yet recruiting | 2023 | NCT06095986 | |
| HK-660S | 23 | Ⅱ | Completed | 2023 | NCT05866809 | |
| Elafibranor | 60 | Ⅱ | Recruiting | 2022 | NCT05627362 | |
| Volixibat | 200 | Ⅱ | Recruiting | 2020 | NCT04663308 | |
| PLN-74809 | 121 | Ⅱ | Completed | 2020 | NCT04480840 | |
| FMT | 58 | Ⅱ | Not yet recruiting | 2024 | NCT06286709 | |
| Vancomycin | 84 | Ⅱ | Recruiting | 2023 | NCT05876182 | |
| BRS201 | 28 | Ⅱ | Recruiting | 2023 | NCT05835505 | |
| CM-101 | 68 | Ⅱ | Not yet recruiting | 2020 | NCT04595825 | |
| EP547 | 58 | Ⅱ | Recruiting | 2022 | NCT05525520 | |
| Rosuvastatin | 256 | Ⅱ | Recruiting | 2023 | NCT05832229 | |
| Curcumin | 15 | Ⅰ/Ⅱ | Completed | 2020 | NCT02978339 | |
| CM-101 | 8 | Ⅰ | Completed | 2023 | NCT06037577 | |
| Rosuvastatin | 15 | Early Ⅰ | Recruiting | 2023 | NCT05912387 | |
| PBC | Cyclosporin A | 89 | Ⅳ | Not yet recruiting | 2020 | NCT04376528 |
| UDCA | 90 | Ⅳ | Recruiting | 2020 | NCT04650243 | |
| UDCA | 90 | Ⅳ | Recruiting | 2020 | NCT04617561 | |
| Babaodan capsule | 60 | Ⅳ | Not yet recruiting | 2024 | NCT06371196 | |
| Fenofibrate | 150 | Ⅲ | Recruiting | 2023 | NCT05751967 | |
| Elafibranor | 161 | Ⅲ | Not yet recruiting | 2020 | NCT04526665 | |
| Elafibranor | 450 | Ⅲ | Recruiting | 2023 | NCT06016842 | |
| Elafibranor | 72 | Ⅲ | Not yet recruiting | 2024 | NCT06383403 | |
| Seladelpar | 192 | Ⅲ | Recruiting | 2023 | NCT06051617 | |
| Seladelpar | 193 | Ⅲ | Completed | 2020 | NCT04620733 | |
| Seladelpar | 150 | Ⅲ | Recruiting | 2023 | NCT06060665 | |
| OCA | 156 | Ⅲ | Recruiting | 2022 | NCT05450887 | |
| OCA | 120 | Ⅲ | Recruiting | 2021 | NCT04956328 | |
| Linerixibat | 230 | Ⅲ | Recruiting | 2021 | NCT04950127 | |
| Linerixibat | 305 | Ⅲ | Recruiting | 2019 | NCT04167358 | |
| Fenofibrate | 184 | Ⅱ/Ⅲ | Recruiting | 2023 | NCT06174402 | |
| Fenofibrate | 104 | Ⅱ/Ⅲ | Recruiting | 2023 | NCT05749822 | |
| Saroglitazar magnesium | 192 | Ⅱ/Ⅲ | Recruiting | 2021 | NCT05133336 | |
| ASC42 | 98 | Ⅱ | Completed | 2022 | NCT05190523 | |
| CS0159 | 75 | Ⅱ | Recruiting | 2023 | NCT05896124 | |
| Volixibat | 180 | Ⅱ | Recruiting | 2021 | NCT05050136 | |
| K-808 | 45 | Ⅱ | Recruiting | 2024 | NCT06247735 | |
| HTD1801 | 24 | Ⅱ | Completed | 2020 | NCT04604652 | |
| EP547 | 58 | Ⅱ | Recruiting | 2022 | NCT05525520 | |
| UC-MSC | 34 | Ⅰ/Ⅱ | Recruiting | 2020 | NCT04522869 | |
| OP-724 | 7 | Ⅰ | Completed | 2019 | NCT04047160 | |
| GSK2330672 | 19 | Ⅰ | Completed | 2019 | NCT04053023 |
-
BA, biliary atresia; PBC, Primary biliary cholangitis; PSC, Primary sclerosing cholangitis; OCA, obeticholic acid; GCSF, granulocyte colony-stimulating factor; BMMC, bone marrow mononuclear stem cell; UC-MSC, umbilical cord-mesenchymal stem cell; FMT, fecal microbiota transplantation; UDCA, ursodeoxycholic acid.
Anti-cholestasis therapy
Bile acid transporters are essential molecules regulating bile acid enterohepatic circulation. Regulating the activity of bile acid transporters alleviates hepatobiliary injury caused by cholestasis, inhibiting cholangiocyte proliferation, preventing or delaying the progression of biliary fibrosis. ASBT is expressed at the ileum and affects reuptake and serum concentration of bile acids. ASBT inhibitors interrupt the bile acid enterohepatic circulation and minimise bile acids accumulation. The results of a randomised controlled trial (RCT) showed that A4250 (Odevixibat, an ASBT inhibitor) efficiently reduced serum levels of bile acids and FGF-19, indicating the potential of ASBT inhibitors for treating PBC and PSC [411]. Baghdasaryan et al. [412] found that A4250 alleviated biliary injury and fibrosis in Mdr2−/− mice, manifesting as reducing serum levels of ALT, ALP and BAs, inhibiting the expressions of TNF-α, Vcam1, Mcp-1, Col1a1 and Colla2, and the proliferation of cholangiocyte. In addition, several pilot studies in PBC patients have shown that Qdevixibat and Linerixibat (ASBT inhibitors) treatment improved the itching symptoms [413]. However, the therapeutic effects of other bile acid transporters remain unclear.
Regulation of bile acid metabolism
UDCA is a hydrophilic and noncytotoxic bile acid approved for treating cholestatic liver disease such as PBC and PSC. In the treatment of PSC, low-dose (10–15 mg/kg/day) and medium-dose (17–23 mg/kg/day) UDCA treatment can improve liver biochemical indicators, but cannot improve long-term prognostic indicators such as liver transplantation risk and 5-year survival rate. However, high-dose UDCA not only does not benefit the user but also increases the chances of requiring liver transplantation and death [47]. In the treatment of PBC, numerous RCTs and meta-analysis studies have shown that UDCA (13–15 mg/kg/day) can improve liver biochemical indicators, delay disease progression, and prolong liver transplant survival time [46, 414]. This dose has been recommended as the standard dosage for PBC treatment [415]. The mechanism of UDCA may be related to cellular protection, anti-inflammatory, and immune regulatory functions. A study showed that UDCA activated PKC-α which in turn inhibited the proliferation and secretion of cholangiocytes [416]. Hatano et al. have demonstrated that oral UDCA alleviated hepatobiliary injury and fibrosis through a pathway independent of bile bicarbonate secretion in the Vil2kd/kd cholestasis mice model [417].
OCA is a semi-synthetic FXR agonist approved as the second-line treatment for PBC [46]. OCA promotes the expression of FGF-19 and further activates FGFR-4/β-KLOTHO pathway, which triggers inhibition of the intracellular pathway of CYP7A1, thereby inhibiting bile acid synthesis and promoting bile acid metabolism. Meanwhile, OCA regulates inflammation, metabolism, and fibrosis through FXR signaling pathway. Clinical studies have shown that OCA combination therapy reduced the levels of ALP and bilirubin and improved long-term outcomes such as liver fibrosis stage, ductular reaction, and bile duct deficiency in the PBC patients with poor response to UDCA [400, 418, 419]. Notably, animal experiments have provided different evidence regarding the role of activating or inhibiting FXR in cholestatic diseases. A study showed that INT-767 (an agonist of TGR5 and FXR) can improve liver inflammation and biliary fibrosis in Mdr2−/− mice [363]. Wagner et al. observed that, FXR knockout reduced the ductal reaction but did not significantly reduce the levels of ALT and ALP in BDL mice [420]. Therefore, further research is needed to elucidate the therapeutic effects and related mechanisms of FXR activation or inhibition in cholangiopathies such as PSC and PBC.
Besides FXR, TGR5 and sphingosine 1-phosphate receptor 2 (S1PR2) have also been reported to be associated with hepatobiliary fibrosis. In the BDL model, the liver condition of TGR5−/− mice significantly deteriorated compared to wild mice, manifested as higher levels of ALT, ALP, IL-6 and TNF-α [421]. Wang et al. demonstrated that S1PR2 expression of cholangiocyte increased in the BDL mouse model, and knocking out S1PR2 or using JTE-013 (an S1PR2 antagonist) reduced BDL-induced hepatobiliary injury and fibrosis [422].
Regulation of inflammation
PPAR agonists show anti-inflammatory and anti-fibrotic effects in cholestatic liver disease. Fenofibrate (PPAR-α agonist) and Bezafibrate (PPAR agonist) are gradually used as second-line treatments for PBC patients with poor UDCA response. Clinical and meta-analysis studies showed that compared with UDCA monotherapy, combined fibrate therapy better improved the levels of ALP, γ-GT, and IgM [399], and reduced the risks of liver transplantation and liver disease-related mortality [47, 423], [424], [425].
Besides fibrate, other PPAR agonists are in clinical trials. Seladelpar (a selective PPAR-β/δ agonist) activates PPAR-δ in hepatocytes, Kupffer cells, and HSCs, inhibiting the synthesis of bile acids and inflammatory cytokines. The phase II clinical trial results showed that Seladelpar continuously improved biochemical indicators such as ALP, ALT, and bilirubin in PBC patients with incomplete or intolerant UDCA response [395]. Among PSC patients treated with Seladelpar 10 mg/day for two years, 79 % achieved the POISE composite endpoint (defined as ALP <1.67 * ULN, ALP decrease ≥15 %, and bilirubin normal), and the overall 2-year non-transplant survival rate was significantly improved [426]. Elafibranor is a dual PPAR-α/δ agonist. Besides the PPAR-δ effects, PPAR-α activation stimulates phospholipid secretion by upregulating MDR3, thereby downregulating the expressions of CYP7A1 and NTCP through phase 1 and 2 enzymes, and reducing inflammation and fibrosis by inhibiting NF-κB [427]. Results of the phase II clinical trial showed that Elafibranor treatment (80 mg/day or 120 mg/day) sustainedly reduced ALP levels in PBC patients with incomplete response or intolerance to UDCA, and improved the proportion of patients achieving the POISE composite endpoint for 12 weeks [394]. Saroglitazar (a dual PPAR-α/γ agonist) promotes the oxidative metabolism of glucose and fatty acids in the liver. The results of a phase II clinical trial showed that Saroglitazar treatment (2 mg/day or 4 mg/day) reduced cholestatic biochemical markers in PBC patients with incomplete response or intolerance to UDCA [428].
Regulation of myofibroblast activation
TGF-β1 is a critical cytokine promoting myofibroblast activation, and inhibiting TGF-β1-induced myofibroblast activation is one of the hot spots in anti-fibrotic therapy [247, 379]. Many studies have confirmed that the TGF-β1/Smads pathway is a target for various compounds to inhibit myofibroblast activation and cholestatic fibrosis, including lactoferrin [429], resveratrol [430], propyleneglycol alginate [248], apigenin [431], physalin D [249], piperlongumine [432], isorhamnetin [433], paclitaxel [434], hesperetin [435], liuweiwuling tablets [436], casticin [437], astragalus total saponins [438], luteolin [439], and allopurinol [440]. TGF-β promotes fibroblast proliferation and activation through the pathways of MET, PPAR-γ, NF-κB and MAPK. Studies have shown that LPS can induce human intrahepatic cholangiocytes to transform into bipolar cells with fibroblast morphology through the TGF-β1 pathway, promoting biliary fibrosis [309].
Epigenetic therapy
NcRNAs promote or inhibit fibrosis by regulating myofibroblasts’ activation and the expression of fibrosis-related genes. Studies have confirmed the potential of targeted fibrosis-related ncRNA therapy in biliary fibrosis. MiRNA-21 can promote enterohepatic dysfunction and fibrosis by regulating host-microbiota crosstalk [374]. MiRNA-21 deletion or miRNA-21 inhibitor treatment can reduce the bone marrow mesenchymal stem cell activation, collagen deposition, and the expression of fibrosis markers such as TGF-β1 and α-SMA [441]. Yang et al. found that exogenous miRNA-29a treatment significantly reduced the ALT and AST levels and the severity of biliary fibrosis in mice [368]. Other studies have shown that lncRNA-H19 deficiency alleviates liver fibrosis in BDL or Mdr2−/− mice, and supplementing cholangiocyte-derived lncRNA-H19 promoted fibrosis in lncRNA-H19 knockout BDL mice [385]. These results indicate that ncRNA has a potential therapeutic role in biliary fibrosis. However, current research evidence mainly comes from animal experiments, and further research is needed for verification. The evidence regarding the association of siRNA and circRNA with fibrosis is lacking.
Immunomodulatory therapy
Immune factors are closely related to the occurrence and development of fibrosis-related cholangiopathies such as PSC, PBC, and IgG4-SC. Limited research provides the potential for immunomodulatory therapy in cholestatic liver injury and fibrosis. Steroid therapy is the standard treatment for IgG4-SC with a response rate of 90 % [442]. In detail, the recommended initial dose of prednisone is 30–40 mg/day for 2–4 weeks, with response evaluation after 2 weeks and then tapered to 5 mg/day (reduce 5 mg/week) over 2–3 months, further reduction or discontinuation of steroid therapy can be considered after 3 years [18, 29, 443]. Notably, tapering off or halting corticosteroid treatment will result in a recurrence of IgG4-SC in approximately 30 % of cases reported in Japan (after 3 years of therapy) and 60 % in Europe (after 3–6 months of therapy) [18]. Upon relapse, reintroduction of corticosteroids alongside other immunosuppressants is indicated [18]. Evidence from meta-analyses and RCTs supports the efficacy of initial combination therapy in substantially reducing the rate of relapse [444], [445], [446]. Rituximab [446, 447] and Azathioprine [444, 445] have demonstrated effectiveness in initiating and sustaining remission, as well as managing recurrences in IgG4-related diseases such as IgG4-SC. A three-year RCT study showed that budesonide (an immunosuppressive agent) combined with UDCA treatment improved liver histological and biochemical indicators in PBC patients with “florid” interface hepatitis, as well as disease activity indicators in recurrent patients [448]. However, the use of budesonide is associated with a higher incidence of serious adverse events. Therefore, current guidelines do not recommend budesonide as a routine treatment for PBC-AIH syndrome but for PBC patients with severe borderline hepatitis [449]. The increase in mast cells around the portal vein area is related to cholangiocyte proliferation and biliary fibrosis in PSC patients [450]. Francis et al. found that compared to wild-type mice, KitW-sh mice with mast cell deficiency significantly reduced BDL-induced biliary injury and fibrosis [192]. Other studies have found that histamine receptor antagonists reduce cholestatic liver injury and fibrosis in both Mdr2−/− and BDL mice [451]. Moreover, Wan et al. found that the expression of neurokinin-1 receptor (NK-1R, an immune-related GPCR) was increased in the liver of PSC patients [452]. Supplementation with L-733060 (an NK-1R antagonist) reduced the expression of fibrosis and senescence genes of HSC and cholangiocyte in Mdr2−/− mouse [452]. Another study found that the cholangiocytes of BDL mice overexpressed NK-1R, and NK-1R−/− mice improved biliary injury and fibrosis after BDL [453]. Therefore, immunomodulatory therapy has excellent potential in treating biliary fibrosis-related cholangiopathies.
Antioxidant therapy
Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX) includes a series of enzymes involved in stress response and is the primary source of ROS in the human body [454]. Low or moderate levels of ROS play essential roles in regulating cell growth, cell signaling, immune response, and autophagy. However, high levels of ROS lead to oxidative stress-induced lipid, protein, and DNA damage, resulting in cell death. NOX1 and NOX4 are highly expressed in endothelial cells, while NOX4 is also expressed in hepatocytes, fibroblasts, and smooth muscle cells. Research has shown that NOX promoted ECM synthesis in HSCs [455], and inhibiting NOX reverses fibrosis in Mdr2−/− mice [266]. Clinical studies have shown that Setanaxib (a selective NOX1/4 inhibitor) improved the levels of ALP and γ-GT in PBC patients. In addition, other studies have reported that the direct application of antioxidants (e.g., vitamin E, silymarin tablets, and anthocyanins) inhibited the activation and proliferation of HSC, and reduced collagen synthesis.
Regulation of microbiota
Intestinal and biliary microbiota dysbiosis has been widely observed in cholangiopathies such as PBC and PSC. Although the specific mechanisms remain unclear, the benefits of regulating microbiota in treating fibrosis-related cholangiopathies have been confirmed [456, 457].
Oral antibiotic therapy (e.g., vancomycin, metronidazole, and minocycline) significantly improves the indicators reflecting disease activity and progression in PSC patients, including clinical symptoms, serum ALP levels, and Mayo score (a long-term prognostic indicator) [458], [459], [460], [461]. The comprehensive results of meta-analysis have shown that antibiotic treatment significantly improved the cholestasis markers in PSC patients, especially vancomycin, which has become an alternative drug for PSC treatment [462]. The mechanism of antibiotic therapy may be related to targeted microorganisms, bile acid metabolism, anti-inflammatory and immune regulation. Vancomycin specifically targets Gram-positive bacteria, such as Clostridium, which converts primary bile acids into secondary bile acids in the distal small intestine and colon [461].
Besides antibiotic therapy, faecal microbiota transplantation (FMT) may benefit biliary fibrosis. FMT reverses gut microbiota imbalance by transferring faecal microbiota from healthy individuals to the intestine of a patient [457]. Previous clinical studies have reported the positive role of FMT in the treatment of Clostridium difficile infection [463, 464] and IBD [465]. Currently, the reports on FMT of cholangiopathies are minimal. Only one pilot study of 10 participants reported that FMT reduced ALP levels in PSC patients with IBD [393]. Therefore, more large-scale studies are needed to verify and explore the efficacy of FMT in biliary fibrosis-related cholangiopathies.
In addition, animal experiments have provided evidence of antifibrotic microbiota therapy. Sterile-fed Mdr2−/− mice exhibited more severe biochemical and histological characteristics of PSC and increased cholangiocyte senescence compared with the conventional-fed, emphasizing the protective role of normal gut microbiota in PSC [466]. Further studies have shown that supplementation with Prevotella copri [467] and Lactobacillus rhamnosus GG [468, 469] significantly improved biliary injury and fibrosis in cholestasis mice. However, another study reported that supplementation with lactic acid bacteria induced a profibrotic inflammatory environment by increasing the translocation of LTA to the liver and upregulating the inflammatory cytokines (e.g., TNF-α, IL-6, and IL-1β) in cholestatic mice [469]. These findings suggest the dual role of microbiota in biliary fibrosis, and clarifying the promotion or inhibition effects of specific microbiota on biliary fibrosis is crucial for microbiota therapy.
Cell therapy
Mesenchymal stem cells (MSC) have been used to treat some benign diseases such as liver cirrhosis, haematological disorders, autoimmune diseases, connective tissue diseases [470], and peripheral nerve injury [471]. Wang et al. have demonstrated that bone marrow MSC transplantation significantly reduced the serum levels of ALP, AMA, and IFN-γ in Poly I: C-induced PBC mice [472]. Lei et al. confirmed that intraperitoneal injection of bone marrow MSC improved the serum levels of AST and ALT, as well as fibrosis markers like TGF-β and type I collagen in the rhesus monkey rotavirus-induced BA model [473]. In addition, many clinical studies have shown that stem cell therapy alleviated liver fibrosis and improved serum indicators of liver function [474]. Notably, cholangiocytes are heterogeneous, small and large cholangiocytes exhibit different stem cell potentials [475]. Previous studies have shown that small cholangiocytes exhibited pluripotent differentiation potential and are differentiated into large cholangiocytes during CCl4-induced injury [475, 476]. Another study showed that small cholangiocyte transplantation reduced ductular reaction and biliary fibrosis in BDL mice, but transplantation of large cholangiocyte had no effect [477]. These findings suggest that small cholangiocytes may have the characteristics of stem cell-like cell populations, and cell therapy may be an effective treatment for biliary fibrosis-related cholangiopathies.
Extracellular vesicle therapy
Extracellular vesicles (EVs) are membrane-bound vesicles secreted by various types of cells, including exosomes and microvesicles, and EVs contain mediators such as DNA, RNA, and proteins. EVs transfer from one cell to another and play essential roles in intercellular communication and bioinformation transmission through proteins, microRNAs, and lncRNAs. Therefore, EV may treat hepatobiliary diseases as a drug or mediator carrier. The unique advantages of EVs in treating liver fibrosis have been initially confirmed. Tan et al. found that stem cell-derived EVs promoted upregulation of the xCT/GPX4 signaling pathway, leading to ferroptosis in HSCs and alleviating liver fibrosis in both LX-2 and CCl4-induced rat models [159]. They also found that EVs isolated from rat bile interacted with cilia of cholangiocyte and inhibit cholangiocyte proliferation in vitro [159]. Another study found that EVs of LPS-induced cholangiocyte promoted normal cholangiocyte inflammation [478]. Chaiyadet et al. found that EVs isolated from hamsters infected with liver flukes promoted normal human cholangiocyte proliferation and IL-6 expression [479]. In addition, studies have reported that miRNA-let7 was delivered to cholangiocytes of Mdr−/− mice through EV injection, thereby exerting a biological effect of inhibiting cholangiocyte proliferation and reducing biliary fibrosis [378]. These findings suggest that both secretory and drug-loaded EVs can regulate ductular reaction, which is one of the characteristics of biliary fibrosis.
Organoids therapy
Organoids, formed through in vitro culture, are three-dimensional cellular aggregates generated by harnessing the specific differentiation and self-assembly properties of stem cells, including embryonic stem cell (ESC), adult stem cell (ASC), and induced pluripotent stem cell (iPSC) [480, 481]. As simplified micro-models of human organs, organoids closely mimic the cellular composition, structure, morphology, and function of real organs, and undergo long-term, stable passaging. Organoids present significant research prospects in fields such as disease modeling, drug screening, developmental biology, precision medicine, and regenerative medicine (Figure 4A) [481]. Organoid injury models play a vital role in the molecular and cellular mechanisms research of tissue damage, repair, and fibrosis. Researchers have successfully simulated the fibrosis process in various organoid injury models, including those of the brain [482], lung [483], heart [484], breast [485], stomach [486], intestine [487], liver [488], biliary tract [489], pancreas [490], and kidney (Figure 4B) [491]. These studies reveal that fibrosis results from interactions among multiple type of cells of injured tissue and aid in understanding the complex cellular interactions and biological functions of mediators in the fibrosis process [492], [493], [494], [495], [496].
Organoids and human disease. (A) Establishment and application of organoids; (B) Established organoid models for simulating the fibrosis process; C. Organoid transplantation for cholangiopathies. Abbreviation: ESC, embryonic stem cell. ASC, adult stem cell. iPSC, induced pluripotent stem cell.
Encouragingly, recent studies provide direct evidence for the therapeutic potential of organoid transplantation in biliary damage repair. Research by Sampaziotis et al. [497] confirmed that features of human cholangiocytes from different regions of biliary tree can be restored in vitro through environmental stimulation. Subsequently, they demonstrated that transplantation of organoids derived from the gallbladder or bile duct enabled repair of injuries throughout the biliary tree in a biliary injury mouse model and a newly developed model for cell transplantation in human organs undergoing ex vivo normothermic perfusion [497]. This result substantiates the feasibility of organoid-based therapies for biliary diseases. Bipotential liver stem cells differentiate into intrahepatic cholangiocytes in vivo [498], and transplanted liver cells formed new intrahepatic bile ducts without pre-existing ductal scaffolds in the Alagille syndrome mouse model [156]. Moreover, cholangiocytes derived from human iPSCs have been proven safe for implantation in the intrahepatic bile ducts of mice [499]. Additionally, with the advent of gene-editing technologies, their integration with organoid technology is poised to bring new breakthroughs in disease mechanism research and cell therapies [500]. Researchers hope to modify organoids using techniques like CRISPR to elucidate tissue changes in patients with or without specific mutations, aiding in identifying disease-causing mutations and mechanisms. Alternatively, gene editing may be used to correct mutations in patient-derived organoids, followed by transplantation of healthy organoids to replace or repair damaged organs, potentially avoiding ethical and immunological issues associated with transplantations (Figure 4C). Although there are currently no human clinical trials of organoid transplantation, organoids will bring significant breakthroughs in the therapy of complex human diseases, with biliary organoid transplantation therapy promising to be a novel treatment method for fibrosis-related cholangiopathies, potentially replacing liver transplantation.
Summary
In this review, we emphasized the importance of biliary fibrosis in hepatobiliary diseases, especially in various cholangiopathies. We summarized the clinical and pathological features of biliary fibrosis, and provided an overview of its etiology. We also introduced the cellular and molecular mechanisms and signaling pathways involved in biliary fibrosis. Lastly, we summarized the therapeutic strategies of biliary fibrosis-related cholangiopathies based on the latest clinical trials and animal experiments. This comprehensive review may offer a valuable reference for mechanism research, drug development, and clinical trials related to biliary fibrosis.
Funding source: National Key Research and Development Program of China
Award Identifier / Grant number: 2022YFC2407405
Funding source: Joint Research Fund Major Project of Gansu Province
Award Identifier / Grant number: 23JRRA1488
Funding source: Medical Innovation and Development Project of Lanzhou University
Award Identifier / Grant number: lzuyxcx-2022-157
Acknowledgments
The authors thank Professor Eske Kvanner Aasvang (Department of Clinical Medicine, Faculty of Health and Medical Sciences, University of Copenhagen, Copenhagen, Denmark) for his suggestions in writing and revising the review.
-
Ethical approval: Not applicable.
-
Informed consent: Not applicable.
-
Author contributions: Conceptualization: Wenbo Meng; Methodology (study design and literature search): Wenbo Meng, Jinyu Zhao, Ping Yue, Ningning Mi, Jinqiu Yuan, Jia Yao, and Yanyan Lin; Writing-original draft: Jinyu Zhao, Ping Yue, Ningning Mi, Matu Li, Wenkang Fu, Xianzhuo Zhang, Long Gao, Mingzhen Bai, Liang Tian, Ningzu Jiang, Yawen Lu, and Haidong Ma, Chunlu Dong, Yong Zhang, Jinduo Zhang, and Yanxian Ren; Writing-review and editing: Wenbo Meng, Hans J Schlitt, Jia Yao, Yanyan Lin, Jinqiu Yuan, Azumi Suzuki, Peng F Wong, Kiyohito Tanaka, Rungsun Rerknimitr, Henrik H Junger, Tan To Cheung, Emmanuel Melloul, Nicolas Demartines, and Joseph W Leung; Visualization (table and figure): Jinyu Zhao, Matu Li, Xianzhuo Zhang, Wenkang Fu, Liang Tian, Jia Yao, and Mingzhen Bai; Supervisor: Wenbo Meng, Jia Yao, Yanyan Lin, and Jinqiu Yuan; Funding acquisition: Wenbo Meng. The authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Competing interests: The authors state no conflict of interest.
-
Research funding: This study was supported by the National Key Research and Development Program of China (2022YFC2407405), the Joint Research Fund Major Project of Gansu Province (23JRRA1488), and the Medical Innovation and Development Project of Lanzhou University (lzuyxcx-2022-157).
-
Data availability: Not applicable.
References
1. Henderson, NC, Rieder, F, Wynn, TA. Fibrosis: from mechanisms to medicines. Nature 2020;587:555–66. https://doi.org/10.1038/s41586-020-2938-9.Search in Google Scholar PubMed PubMed Central
2. Pakshir, P, Hinz, B. The big five in fibrosis: macrophages, myofibroblasts, matrix, mechanics, and miscommunication. Matrix Biol 2018;68–69:81–93. https://doi.org/10.1016/j.matbio.2018.01.019.Search in Google Scholar PubMed
3. Wynn, TA. Cellular and molecular mechanisms of fibrosis. J Pathol 2008;214:199–210. https://doi.org/10.1002/path.2277.Search in Google Scholar PubMed PubMed Central
4. Rockey, DC, Bell, PD, Hill, JA. Fibrosis--a common pathway to organ injury and failure. N Engl J Med 2015;372:1138–49. https://doi.org/10.1056/nejmra1300575.Search in Google Scholar
5. Noble, PW, Barkauskas, CE, Jiang, D. Pulmonary fibrosis: patterns and perpetrators. J Clin Invest 2012;122:2756–62. https://doi.org/10.1172/jci60323.Search in Google Scholar PubMed PubMed Central
6. Nagalingam, RS, Chattopadhyaya, S, Al-Hattab, DS, Cheung, DYC, Schwartz, LY, Jana, S, et al.. Scleraxis and fibrosis in the pressure-overloaded heart. Eur Heart J 2022;43:4739–50. https://doi.org/10.1093/eurheartj/ehac362.Search in Google Scholar PubMed
7. Bataller, R, Brenner, DA. Liver fibrosis. J Clin Invest 2005;115:209–18. https://doi.org/10.1172/jci24282.Search in Google Scholar PubMed PubMed Central
8. Huang, R, Fu, P, Ma, L. Kidney fibrosis: from mechanisms to therapeutic medicines. Signal Transduct Targeted Ther 2023;8:129. https://doi.org/10.1038/s41392-023-01379-7.Search in Google Scholar PubMed PubMed Central
9. Katabathina, VS, Flaherty, EM, Dasyam, AK, Menias, CO, Riddle, ND, Lath, N, et al.. “Biliary diseases with pancreatic counterparts”: cross-sectional imaging findings. Radiographics 2016;36:374–92. https://doi.org/10.1148/rg.2016150071.Search in Google Scholar PubMed
10. Zahr, AA, Salama, ME, Carreau, N, Tremblay, D, Verstovsek, S, Mesa, R, et al.. Bone marrow fibrosis in myelofibrosis: pathogenesis, prognosis and targeted strategies. Haematologica 2016;101:660–71. https://doi.org/10.3324/haematol.2015.141283.Search in Google Scholar PubMed PubMed Central
11. Lenti, MV, Di Sabatino, A. Intestinal fibrosis. Mol Aspect Med 2019;65:100–9. https://doi.org/10.1016/j.mam.2018.10.003.Search in Google Scholar PubMed
12. D’Alessio, S, Ungaro, F, Noviello, D, Lovisa, S, Peyrin-Biroulet, L, Danese, S. Revisiting fibrosis in inflammatory bowel disease: the gut thickens. Nat Rev Gastroenterol Hepatol 2022;19:169–84. https://doi.org/10.1038/s41575-021-00543-0.Search in Google Scholar PubMed
13. Zhao, M, Wang, L, Wang, M, Zhou, S, Lu, Y, Cui, H, et al.. Targeting fibrosis, mechanisms and cilinical trials. Signal Transduct Targeted Ther 2022;7:206. https://doi.org/10.1038/s41392-022-01070-3.Search in Google Scholar PubMed PubMed Central
14. Ando, H, Inomata, Y, Iwanaka, T, Kuroda, T, Nio, M, Matsui, A, et al.. Clinical practice guidelines for biliary atresia in Japan: a secondary publication of the abbreviated version translated into English. J Hepatobiliary Pancreat Sci 2021;28:55–61. https://doi.org/10.1002/jhbp.816.Search in Google Scholar PubMed
15. Venkatesh, SK, Welle, CL, Miller, FH, Jhaveri, K, Ringe, KI, Eaton, JE, et al.. Reporting standards for primary sclerosing cholangitis using MRI and MR cholangiopancreatography: guidelines from MR working group of the international primary sclerosing cholangitis study group. Eur Radiol 2022;32:923–37. https://doi.org/10.1007/s00330-021-08147-7.Search in Google Scholar PubMed
16. Möller, K, Braden, B, Culver, EL, Jenssen, C, Zadeh, ES, Alhyari, A, et al.. Secondary sclerosing cholangitis and IgG4-sclerosing cholangitis - a review of cholangiographic and ultrasound imaging. Endosc Ultrasound 2023;12:181–99. https://doi.org/10.4103/eus-d-22-00208.Search in Google Scholar PubMed PubMed Central
17. Zhang, Y, Zheng, T, Huang, Z, Song, B. CT and MR imaging of primary biliary cholangitis: a pictorial review. Insights Imag 2023;14:180. https://doi.org/10.1186/s13244-023-01517-3.Search in Google Scholar PubMed PubMed Central
18. Löhr, JM, Beuers, U, Vujasinovic, M, Alvaro, D, Frøkjær, JB, Buttgereit, F, et al.. European Guideline on IgG4-related digestive disease - UEG and SGF evidence-based recommendations. United European Gastroenterol J 2020;8:637–66. https://doi.org/10.1177/2050640620934911.Search in Google Scholar PubMed PubMed Central
19. Junger, HH, Schlitt, HJ, Geissler, EK, Fichtner-Feigl, S, Brunner, SM. Bile duct regeneration and immune response by passenger lymphocytes signals biliary recovery versus complications after liver transplantation. Liver Transplant 2017;23:1422–32. https://doi.org/10.1002/lt.24836.Search in Google Scholar PubMed
20. Friedman, SL, Pinzani, M. Hepatic fibrosis 2022: unmet needs and a blueprint for the future. Hepatology 2022;75:473–88. https://doi.org/10.1002/hep.32285.Search in Google Scholar PubMed
21. Parola, M, Pinzani, M. Liver fibrosis: pathophysiology, pathogenetic targets and clinical issues. Mol Aspect Med 2019;65:37–55. https://doi.org/10.1016/j.mam.2018.09.002.Search in Google Scholar PubMed
22. Chen, G, Jiang, J, Tang, Y, Zhu, Y, Xiao, X, Wang, J, et al.. Evidence-based practice guidelines on the diagnosis and treatment of biliary atresia. Chin J Evid Base Pediatr 2022;17:245–59.Search in Google Scholar
23. Vij, M, Rela, M. Biliary atresia: pathology, etiology and pathogenesis. Future Sci OA 2020;6:Fso466. https://doi.org/10.2144/fsoa-2019-0153.Search in Google Scholar PubMed PubMed Central
24. You, H, Ma, X, Efe, C, Wang, G, Jeong, SH, Abe, K, et al.. APASL clinical practice guidance: the diagnosis and management of patients with primary biliary cholangitis. Hepatol Int 2022;16:1–23. https://doi.org/10.1007/s12072-021-10276-6.Search in Google Scholar PubMed PubMed Central
25. You, H, Duan, W, Li, S, Lv, T, Chen, S, Lu, L, et al.. Guidelines on the diagnosis and management of primary biliary cholangitis (2021). J Clin Transl Hepatol 2023;11:736–46. https://doi.org/10.14218/JCTH.2022.00347.Search in Google Scholar PubMed PubMed Central
26. Chapman, MH, Thorburn, D, Hirschfield, GM, Webster, GGJ, Rushbrook, SM, Alexander, G, et al.. British Society of Gastroenterology and UK-PSC guidelines for the diagnosis and management of primary sclerosing cholangitis. Gut 2019;68:1356–78. https://doi.org/10.1136/gutjnl-2018-317993.Search in Google Scholar PubMed PubMed Central
27. Tow, CY, Chung, E, Kaul, B, Bhalla, A, Fortune, BE. Diagnostic tests in primary sclerosing cholangitis: serology, elastography, imaging, and histology. Clin Liver Dis 2024;28:157–69. https://doi.org/10.1016/j.cld.2023.07.007.Search in Google Scholar PubMed
28. Liver, EAS, Beuers, U, Bergquist, A, Karlsen, TH, Levy, C, Samyn, M, et al.. EASL clinical practice guidelines on sclerosing cholangitis. J Hepatol 2022;77:761–806. https://doi.org/10.1016/j.jhep.2022.05.011.Search in Google Scholar PubMed
29. Kamisawa, T, Nakazawa, T, Tazuma, S, Zen, Y, Tanaka, A, Ohara, H, et al.. Clinical practice guidelines for IgG4-related sclerosing cholangitis. J Hepatobiliary Pancreat Sci 2019;26:9–42. https://doi.org/10.1002/jhbp.596.Search in Google Scholar PubMed PubMed Central
30. Feranchak, AP. Hepatobiliary complications of cystic fibrosis. Curr Gastroenterol Rep 2004;6:231–9. https://doi.org/10.1007/s11894-004-0013-6.Search in Google Scholar PubMed
31. Brown, ZJ, Baghdadi, A, Kamel, I, Labiner, HE, Hewitt, DB, Pawlik, TM. Diagnosis and management of choledochal cysts. HPB 2023;25:14–25. https://doi.org/10.1016/j.hpb.2022.09.010.Search in Google Scholar PubMed
32. Lasagni, A, Cadamuro, M, Morana, G, Fabris, L, Strazzabosco, M. Fibrocystic liver disease: novel concepts and translational perspectives. Transl Gastroenterol Hepatol 2021;6:26. https://doi.org/10.21037/tgh-2020-04.Search in Google Scholar PubMed PubMed Central
33. Ruemmele, P, Hofstaedter, F, Gelbmann, CM. Secondary sclerosing cholangitis. Nat Rev Gastroenterol Hepatol 2009;6:287–95. https://doi.org/10.1038/nrgastro.2009.46.Search in Google Scholar PubMed
34. Lv, T, Chen, S, Li, M, Zhang, D, Kong, Y, Jia, J. Regional variation and temporal trend of primary biliary cholangitis epidemiology: a systematic review and meta-analysis. J Gastroenterol Hepatol 2021;36:1423–34. https://doi.org/10.1111/jgh.15329.Search in Google Scholar PubMed
35. Wojcicki, M, Milkiewicz, P, Silva, M. Biliary tract complications after liver transplantation: a review. Dig Surg 2008;25:245–57. https://doi.org/10.1159/000144653.Search in Google Scholar PubMed
36. Heidenhain, C, Pratschke, J, Puhl, G, Neumann, U, Pascher, A, Veltzke-Schlieker, W, et al.. Incidence of and risk factors for ischemic-type biliary lesions following orthotopic liver transplantation. Transpl Int 2010;23:14–22. https://doi.org/10.1111/j.1432-2277.2009.00947.x.Search in Google Scholar PubMed
37. Pollock, G, Minuk, GY. Diagnostic considerations for cholestatic liver disease. J Gastroenterol Hepatol 2017;32:1303–9. https://doi.org/10.1111/jgh.13738.Search in Google Scholar PubMed
38. Herta, T, Verheij, J, Beuers, U. IgG4-assoziierte Cholangitis – klinische Präsentation eines lange übersehenen Krankheitsbildes [IgG4-associated cholangitis - clinical presentation of an overlooked disease entity]. Internist 2018;59:560–6. https://doi.org/10.1007/s00108-018-0431-4.Search in Google Scholar PubMed
39. Brahee, DD, Lampl, BS. Neonatal diagnosis of biliary atresia: a practical review and update. Pediatr Radiol 2022;52:685–92. https://doi.org/10.1007/s00247-021-05148-y.Search in Google Scholar PubMed
40. Khoshpouri, P, Habibabadi, RR, Hazhirkarzar, B, Ameli, S, Ghadimi, M, Ghasabeh, MA, et al.. Imaging features of primary sclerosing cholangitis: from diagnosis to liver transplant follow-up. Radiographics 2019;39:1938–64. https://doi.org/10.1148/rg.2019180213.Search in Google Scholar PubMed
41. Morgan, MA, Khot, R, Sundaram, KM, Ludwig, DR, Nair, RT, Mittal, PK, et al.. Primary sclerosing cholangitis: review for radiologists. Abdom Radiol 2023;48:136–50. https://doi.org/10.1007/s00261-022-03655-6.Search in Google Scholar PubMed PubMed Central
42. Nguyen, DL, Juran, BD, Lazaridis, KN. Primary biliary cirrhosis. Best Pract Res Clin Gastroenterol 2010;24:647–54. https://doi.org/10.1016/j.bpg.2010.07.006.Search in Google Scholar PubMed PubMed Central
43. Kovač, JD, Milovanović, T. MRI periportal halo sign" in primary biliary cirrhosis. Abdom Radiol 2023;48:1556–7. https://doi.org/10.1007/s00261-023-03859-4.Search in Google Scholar PubMed
44. Pouw, RE, Barret, M, Biermann, K, Bisschops, R, Czakó, L, Gecse, KB, et al.. Endoscopic tissue sampling - Part 1: upper gastrointestinal and hepatopancreatobiliary tracts. European society of gastrointestinal endoscopy (ESGE) guideline. Endoscopy 2021;53:1174–88. https://doi.org/10.1055/a-1611-5091.Search in Google Scholar PubMed
45. Mauro, A, Mazza, S, Scalvini, D, Lusetti, F, Bardone, M, Quaretti, P, et al.. The role of cholangioscopy in biliary diseases. Diagnostics 2023;13:2933. https://doi.org/10.3390/diagnostics13182933.Search in Google Scholar PubMed PubMed Central
46. Hepatology CSo. Guidelines on the diagnosis and management of primary biliary cholangitis (2021). Chin J Intern Med 2021;60:1024–37. https://doi.org/10.3760/cma.j.cn112138-20211112-00794.Search in Google Scholar PubMed
47. Hepatology CSo. Guidelines on the diagnosis and management of primary sclerosing cholangitis (2021). Chin J Intern Med 2021;60:1050–74. https://doi.org/10.3760/cma.j.cn112138-20211109-00786.Search in Google Scholar PubMed
48. Sadeghi, A, Mohamadnejad, M, Islami, F, Keshtkar, A, Biglari, M, Malekzadeh, R, et al.. Diagnostic yield of EUS-guided FNA for malignant biliary stricture: a systematic review and meta-analysis. Gastrointest Endosc 2016;83:290–8.e1. https://doi.org/10.1016/j.gie.2015.09.024.Search in Google Scholar PubMed
49. Jin, Z. The application of endoscopic ultrasonography in the digestive diseases. Chin J Gastroenterol Hepatol 2009;18:5–9.Search in Google Scholar
50. Deng, W, Jiang, H, Qin, S. Application progress of contrast-enhanced endoscopic ultrasonography in the diagnosis and treatment of digestive system diseases. Guangxi Med J 2017;39:1403–5.Search in Google Scholar
51. Farrell, RJ, Agarwal, B, Brandwein, SL, Underhill, J, Chuttani, R, Pleskow, DK. Intraductal US is a useful adjunct to ERCP for distinguishing malignant from benign biliary strictures. Gastrointest Endosc 2002;56:681–7. https://doi.org/10.1016/s0016-5107(02)70117-x.Search in Google Scholar
52. Heinzow, HS, Kammerer, S, Rammes, C, Wessling, J, Domagk, D, Meister, T. Comparative analysis of ERCP, IDUS, EUS and CT in predicting malignant bile duct strictures. World J Gastroenterol 2014;20:10495–503. https://doi.org/10.3748/wjg.v20.i30.10495.Search in Google Scholar PubMed PubMed Central
53. Della-Torre, E, Feeney, E, Deshpande, V, Mattoo, H, Mahajan, V, Kulikova, M, et al.. B-cell depletion attenuates serological biomarkers of fibrosis and myofibroblast activation in IgG4-related disease. Ann Rheum Dis 2015;74:2236–43. https://doi.org/10.1136/annrheumdis-2014-205799.Search in Google Scholar PubMed PubMed Central
54. Mayo, MJ, Parkes, J, Adams-Huet, B, Combes, B, Mills, AS, Markin, RS, et al.. Prediction of clinical outcomes in primary biliary cirrhosis by serum enhanced liver fibrosis assay. Hepatology 2008;48:1549–57. https://doi.org/10.1002/hep.22517.Search in Google Scholar PubMed PubMed Central
55. Fossdal, G, Mjelle, AB, Wiencke, K, Bjørk, I, Gilja, OH, Folseraas, T, et al.. Fluctuating biomarkers in primary sclerosing cholangitis: a longitudinal comparison of alkaline phosphatase, liver stiffness, and ELF. JHEP Rep 2021;3:100328. https://doi.org/10.1016/j.jhepr.2021.100328.Search in Google Scholar PubMed PubMed Central
56. Sato, K, Glaser, S, Kennedy, L, Liangpunsakul, S, Meng, F, Francis, H, et al.. Preclinical insights into cholangiopathies: disease modeling and emerging therapeutic targets. Expert Opin Ther Targets 2019;23:461–72. https://doi.org/10.1080/14728222.2019.1608950.Search in Google Scholar PubMed PubMed Central
57. Liu, JZ, Hov, JR, Folseraas, T, Ellinghaus, E, Rushbrook, SM, Doncheva, NT, et al.. Dense genotyping of immune-related disease regions identifies nine new risk loci for primary sclerosing cholangitis. Nat Genet 2013;45:670–5. https://doi.org/10.1038/ng.2616.Search in Google Scholar PubMed PubMed Central
58. Quelhas, P, Cerski, C, Dos Santos, JL. Update on etiology and pathogenesis of biliary atresia. Curr Pediatr Rev 2022;19:48–67. https://doi.org/10.2174/1573396318666220510130259.Search in Google Scholar PubMed
59. Fawaz, R, Baumann, U, Ekong, U, Fischler, B, Hadzic, N, Mack, CL, et al.. Guideline for the evaluation of cholestatic jaundice in infants: Joint recommendations of the north American society for pediatric gastroenterology, hepatology, and nutrition and the European society for pediatric gastroenterology, hepatology, and nutrition. J Pediatr Gastroenterol Nutr 2017;64:154–68. https://doi.org/10.1097/mpg.0000000000001334.Search in Google Scholar PubMed
60. Le Roy, B, Gagnière, J, Filaire, L, Fontarensky, M, Hordonneau, C, Buc, E. Pancreaticobiliary maljunction and choledochal cysts: from embryogenesis to therapeutics aspects. Surg Radiol Anat 2016;38:1053–60. https://doi.org/10.1007/s00276-016-1669-y.Search in Google Scholar PubMed
61. Kuraishi, Y, Uehara, T, Muraki, T, Iwaya, M, Kinugawa, Y, Nakajima, T, et al.. Impact of DNA double-strand breaks on pancreaticobiliary maljunction carcinogenesis. Diagn Pathol 2021;16:72. https://doi.org/10.1186/s13000-021-01132-0.Search in Google Scholar PubMed PubMed Central
62. Khan, MZ, Kichloo, A, El-Amir, Z, Shah Zaib, M, Wani, F. Caroli disease: a presentation of acute pancreatitis and cholangitis. Cureus 2020;12:e9135. https://doi.org/10.7759/cureus.9135.Search in Google Scholar PubMed PubMed Central
63. Zimmerman, KA, Song, CJ, Aloria, EJG, Li, Z, Zhou, J, Bland, SJ, et al.. Early infiltrating macrophage subtype correlates with late-stage phenotypic outcome in a mouse model of hepatorenal fibrocystic disease. Lab Invest 2021;101:1382–93. https://doi.org/10.1038/s41374-021-00627-0.Search in Google Scholar PubMed PubMed Central
64. Meyerholz, DK, Leidinger, MR, Adam Goeken, J, Businga, TR, Vizuett, S, Akers, A, et al.. Immunohistochemical detection of MUC5AC and MUC5B mucins in ferrets. BMC Res Notes 2023;16:111. https://doi.org/10.1186/s13104-023-06388-x.Search in Google Scholar PubMed PubMed Central
65. Dana, J, Debray, D, Beaufrère, A, Hillaire, S, Fabre, M, Reinhold, C, et al.. Cystic fibrosis-related liver disease: clinical presentations, diagnostic and monitoring approaches in the era of CFTR modulator therapies. J Hepatol 2022;76:420–34. https://doi.org/10.1016/j.jhep.2021.09.042.Search in Google Scholar PubMed
66. Gulamhusein, AF, Juran, BD, Lazaridis, KN. Genome-wide association studies in primary biliary cirrhosis. Semin Liver Dis 2015;35:392–401. https://doi.org/10.1055/s-0035-1567831.Search in Google Scholar PubMed PubMed Central
67. Carbone, M, Lleo, A, Sandford, RN, Invernizzi, P. Implications of genome-wide association studies in novel therapeutics in primary biliary cirrhosis. Eur J Immunol 2014;44:945–54. https://doi.org/10.1002/eji.201344270.Search in Google Scholar PubMed PubMed Central
68. Gulamhusein, AF, Hirschfield, GM. Primary biliary cholangitis: pathogenesis and therapeutic opportunities. Nat Rev Gastroenterol Hepatol 2020;17:93–110. https://doi.org/10.1038/s41575-019-0226-7.Search in Google Scholar PubMed
69. Wang, C, Zheng, X, Jiang, P, Tang, R, Gong, Y, Dai, Y, et al.. Genome-wide association studies of specific antinuclear autoantibody subphenotypes in primary biliary cholangitis. Hepatology 2019;70:294–307. https://doi.org/10.1002/hep.30604.Search in Google Scholar PubMed PubMed Central
70. Hirschfield, GM, Gershwin, ME. The immunobiology and pathophysiology of primary biliary cirrhosis. Annu Rev Pathol 2013;8:303–30. https://doi.org/10.1146/annurev-pathol-020712-164014.Search in Google Scholar PubMed
71. Diseases, NCRCI. Expert consensus on the diagnosis and treatment of intrahepatic cholestasis (2021 edition). Chin J Hepatol 2021;14:401–12.Search in Google Scholar
72. Bowlus, CL, Arrivé, L, Bergquist, A, Deneau, M, Forman, L, Ilyas, SI, et al.. AASLD practice guidance on primary sclerosing cholangitis and cholangiocarcinoma. Hepatology 2023;77:659–702. https://doi.org/10.1002/hep.32771.Search in Google Scholar PubMed
73. Berglin, L, Bergquist, A, Johansson, H, Glaumann, H, Jorns, C, Lunemann, S, et al.. In situ characterization of intrahepatic non-parenchymal cells in PSC reveals phenotypic patterns associated with disease severity. PLoS One 2014;9:e105375. https://doi.org/10.1371/journal.pone.0105375.Search in Google Scholar PubMed PubMed Central
74. Gwela, A, Siddhanathi, P, Chapman, RW, Travis, S, Powrie, F, Arancibia-Cárcamo, CV, et al.. Th1 and innate lymphoid cells accumulate in primary sclerosing cholangitis-associated inflammatory bowel disease. J Crohns Colitis 2017;11:1124–34. https://doi.org/10.1093/ecco-jcc/jjx050.Search in Google Scholar PubMed PubMed Central
75. Koda, S, Zhu, XQ, Zheng, KY, Yan, C. Molecular mechanisms of Clonorchis sinensis-host interactions and implications for vaccine development. Front Cell Dev Biol 2021;9:781768. https://doi.org/10.3389/fcell.2021.781768.Search in Google Scholar PubMed PubMed Central
76. Wang, Y, Zhang, X, Wang, X, Zhang, N, Yu, Y, Gong, P, et al.. Clonorchis sinensis aggravates biliary fibrosis through promoting IL-6 production via toll-like receptor 2-mediated AKT and p38 signal pathways. PLoS Neglected Trop Dis 2023;17:e0011062. https://doi.org/10.1371/journal.pntd.0011062.Search in Google Scholar PubMed PubMed Central
77. Yan, C, Wu, J, Xu, N, Li, J, Zhou, QY, Yang, HM, et al.. TLR4 deficiency exacerbates biliary injuries and peribiliary fibrosis caused by Clonorchis sinensis in a resistant mouse strain. Front Cell Infect Microbiol 2020;10:526997. https://doi.org/10.3389/fcimb.2020.526997.Search in Google Scholar PubMed PubMed Central
78. Khuroo, MS, Rather, AA, Khuroo, NS, Khuroo, MS. Hepatobiliary and pancreatic ascariasis. World J Gastroenterol 2016;22:7507–17. https://doi.org/10.3748/wjg.v22.i33.7507.Search in Google Scholar PubMed PubMed Central
79. Yamashita, M, Adachi, T, Ono, S, Yoshino, K, Imamura, H, Matsushima, H, et al.. Helicobacter bilis infection induces oxidative stress in and enhances the proliferation of human cholangiocytes. Helicobacter 2022;27:e12908. https://doi.org/10.1111/hel.12908.Search in Google Scholar PubMed
80. Zigmond, E, Zecher, BF, Bartels, AL, Ziv-Baran, T, Rösch, T, Schachschal, G, et al.. Bile duct colonization with Enterococcus sp. associates with disease progression in primary sclerosing cholangitis. Clin Gastroenterol Hepatol 2023;21:1223–32.e3. https://doi.org/10.1016/j.cgh.2022.09.006.Search in Google Scholar PubMed
81. Bruneau, A, Hundertmark, J, Guillot, A, Tacke, F. Molecular and cellular mediators of the gut-liver Axis in the progression of liver diseases. Front Med 2021;8:725390. https://doi.org/10.3389/fmed.2021.725390.Search in Google Scholar PubMed PubMed Central
82. De Angelis, C, Mangone, M, Bianchi, M, Saracco, G, Repici, A, Rizzetto, M, et al.. An update on AIDS-related cholangiopathy. Minerva Gastroenterol Dietol 2009;55:79–82.Search in Google Scholar
83. Hidalgo-Tenorio, C, Blasco-Morente, G. Sclerosing cholangitis by cytomegalovirus in highly active antiretroviral therapy era. Rev Esp Enferm Dig 2013;105:557–60. https://doi.org/10.4321/s1130-01082013000900009.Search in Google Scholar PubMed
84. Hartl, L, Haslinger, K, Angerer, M, Semmler, G, Schneeweiss-Gleixner, M, Jachs, M, et al.. Progressive cholestasis and associated sclerosing cholangitis are frequent complications of COVID-19 in patients with chronic liver disease. Hepatology 2022;76:1563–75. https://doi.org/10.1002/hep.32582.Search in Google Scholar PubMed PubMed Central
85. Leonhardt, S, Jürgensen, C, Frohme, J, Grajecki, D, Adler, A, Sigal, M, et al.. Hepatobiliary long-term consequences of COVID-19: dramatically increased rate of secondary sclerosing cholangitis in critically ill COVID-19 patients. Hepatol Int 2023;17:1610–25. https://doi.org/10.1007/s12072-023-10521-0.Search in Google Scholar PubMed PubMed Central
86. Dobreva, I, Karagyozov, P. Drug-induced bile duct injury - a short review. Curr Drug Metabol 2020;21:256–9. https://doi.org/10.2174/1389200221666200420100129.Search in Google Scholar PubMed
87. Ma, MX, Jayasekeran, V, Chong, AK. Benign biliary strictures: prevalence, impact, and management strategies. Clin Exp Gastroenterol 2019;12:83–92. https://doi.org/10.2147/ceg.s165016.Search in Google Scholar PubMed PubMed Central
88. Sandrasegaran, K, Alazmi, WM, Tann, M, Fogel, EL, McHenry, L, Lehman, GA. Chemotherapy-induced sclerosing cholangitis. Clin Radiol 2006;61:670–8. https://doi.org/10.1016/j.crad.2006.02.013.Search in Google Scholar PubMed
89. Horsley-Silva, JL, Dow, EN, Menias, CO, Smith, ML, Carballido, EM, Lindor, KD, et al.. Docetaxel induced sclerosing cholangitis. Dig Dis Sci 2015;60:3814–6. https://doi.org/10.1007/s10620-015-3914-3.Search in Google Scholar PubMed
90. Kusakabe, A, Ohkawa, K, Fukutake, N, Sakakibara, M, Imai, T, Abe, Y, et al.. Chemotherapy-induced sclerosing cholangitis caused by systemic chemotherapy. ACG Case Rep J 2019;6:e00136. https://doi.org/10.14309/crj.0000000000000136.Search in Google Scholar PubMed PubMed Central
91. Kawabata, H, Honda, M, Kawamura, T, Kori, Y, Uenoyama, Y, Kawaguchi, Y, et al.. [A case of primary sclerosing cholangitis (PSC) complicated with acute inflammatory demyelinating polyradiculoneuropathy]. Nihon Shokakibyo Gakkai Zasshi 2002;99:1503–7.Search in Google Scholar
92. Pi, B, Wang, J, Tong, Y, Yang, Q, Lv, F, Yu, Y. Immune-related cholangitis induced by immune checkpoint inhibitors: a systematic review of clinical features and management. Eur J Gastroenterol Hepatol 2021;33(1S Suppl 1):e858–67. https://doi.org/10.1097/meg.0000000000002280.Search in Google Scholar PubMed PubMed Central
93. Koya, Y, Shibata, M, Shinohara, N, Nebuya, S, Oe, S, Honma, Y, et al.. Secondary sclerosing cholangitis with hemobilia induced by pembrolizumab: case report and review of published work. Hepatol Res 2019;49:950–6. https://doi.org/10.1111/hepr.13329.Search in Google Scholar PubMed
94. Kawakami, H, Tanizaki, J, Tanaka, K, Haratani, K, Hayashi, H, Takeda, M, et al.. Imaging and clinicopathological features of nivolumab-related cholangitis in patients with non-small cell lung cancer. Invest N Drugs 2017;35:529–36. https://doi.org/10.1007/s10637-017-0453-0.Search in Google Scholar PubMed
95. Kashima, J, Okuma, Y, Shimizuguchi, R, Chiba, K. Bile duct obstruction in a patient treated with nivolumab as second-line chemotherapy for advanced non-small-cell lung cancer: a case report. Cancer Immunol Immunother 2018;67:61–5. https://doi.org/10.1007/s00262-017-2062-3.Search in Google Scholar PubMed PubMed Central
96. Nakano, R, Shiomi, H, Fujiwara, A, Yoshihara, K, Yoshioka, R, Kawata, S, et al.. Clinical characteristics of ICI-related pancreatitis and cholangitis including radiographic and endoscopic findings. Healthcare 2022;10. https://doi.org/10.3390/healthcare10050763.Search in Google Scholar PubMed PubMed Central
97. Zen, Y, Chen, YY, Jeng, YM, Tsai, HW, Yeh, MM. Immune-related adverse reactions in the hepatobiliary system: second-generation check-point inhibitors highlight diverse histological changes. Histopathology 2020;76:470–80. https://doi.org/10.1111/his.14000.Search in Google Scholar PubMed
98. Turkish, A, Luo, JJ, Lefkowitch, JH. Ketamine abuse, biliary tract disease, and secondary sclerosing cholangitis. Hepatology 2013;58:825–7. https://doi.org/10.1002/hep.26459.Search in Google Scholar PubMed
99. Zeng, J, Luo, F, Lin, Z, Chen, Y, Wang, X, Song, Y. Rash and cholestatic liver injury caused by methimazole in a woman with Turner syndrome and Graves’s disease: a case report and literature review. BMC Endocr Disord 2021;21:179. https://doi.org/10.1186/s12902-021-00819-1.Search in Google Scholar PubMed PubMed Central
100. Ahmad, J, Rossi, S, Rodgers, SK, Ghabril, M, Fontana, RJ, Stolz, A, et al.. Sclerosing cholangitis-like changes on magnetic resonance cholangiography in patients with drug induced liver injury. Clin Gastroenterol Hepatol 2019;17:789–90. https://doi.org/10.1016/j.cgh.2018.06.035.Search in Google Scholar PubMed PubMed Central
101. Rymbai, ML, Paul, A, M, JA, Anantrao, AS, John, R, Simon, B, et al.. Post cholecystectomy benign biliary stricture-isolated hepatic duct stricture: a proposed modification of the BISMUTH classification. ANZ J Surg 2023;93:1306–13. https://doi.org/10.1111/ans.18281.Search in Google Scholar PubMed
102. Reinsoo, A, Kirsimägi, Ü, Kibuspuu, L, Košeleva, K, Lepner, U, Talving, P. Bile duct injuries during laparoscopic cholecystectomies: an 11-year population-based study. Eur J Trauma Emerg Surg 2023;49:2269–76. https://doi.org/10.1007/s00068-022-02190-9.Search in Google Scholar PubMed
103. Anand, U, Kumar, R, Priyadarshi, RN, Kumar, M, Kumar, R, Ahmed, N, et al.. Analysis of outcomes of biliary reconstruction after post-cholecystectomy bile duct injuries. ANZ J Surg 2021;91:1542–8. https://doi.org/10.1111/ans.17047.Search in Google Scholar PubMed
104. Kimura, K, Yoshizumi, T, Kudo, K, Oh, K, Kurihara, T, Toshima, T, et al.. Intractable biliary strictures after living donor liver transplantation: a case series. Transplant Proc 2021;53:1726–30. https://doi.org/10.1016/j.transproceed.2021.04.015.Search in Google Scholar PubMed
105. Zhao, J, Tian, L, Xia, B, Mi, N, He, Q, Yang, M, et al.. Cholecystectomy is associated with a higher risk of irritable bowel syndrome in the UK Biobank: a prospective cohort study. Front Pharmacol 2023;14:1244563. https://doi.org/10.3389/fphar.2023.1244563.Search in Google Scholar PubMed PubMed Central
106. Manzoor, UHM, Hassan Luck, N, Ali Tasneem, A, Mudassir Laeeq, S, Mandhwani, R, Hanif, FM, et al.. Safety and efficacy of extracorporeal shock wave lithotripsy for difficult-to-retrieve common bile duct stones: a ten-year experience. J Transl Int Med 2020;8:159–64. https://doi.org/10.2478/jtim-2020-0025.Search in Google Scholar PubMed PubMed Central
107. Chao, J, Cui, S, Liu, C, Liu, S, Liu, S, Han, Y, et al.. Detection of early cytokine storm in patients with septic shock after abdominal surgery. J Transl Int Med 2020;8:91–8. https://doi.org/10.2478/jtim-2020-0014.Search in Google Scholar PubMed PubMed Central
108. Deltenre, P, Valla, DC. Ischemic cholangiopathy. J Hepatol 2006;44:806–17. https://doi.org/10.1016/j.jhep.2006.01.009.Search in Google Scholar PubMed
109. Deltenre, P, Valla, DC. Ischemic cholangiopathy. Semin Liver Dis 2008;28:235–46. https://doi.org/10.1055/s-0028-1085092.Search in Google Scholar PubMed
110. Ly, M, Lau, NS, McKenzie, C, Kench, JG, Seyfi, D, Majumdar, A, et al.. Histological assessment of the bile duct before liver transplantation: does the bile duct injury score predict biliary strictures? J Clin Med 2023;12. https://doi.org/10.3390/jcm12216793.Search in Google Scholar PubMed PubMed Central
111. Martins, P, Verdelho Machado, M. Secondary sclerosing cholangitis in critically ill patients: an underdiagnosed entity. GE Port J Gastroenterol 2020;27:103–14. https://doi.org/10.1159/000501405.Search in Google Scholar PubMed PubMed Central
112. Leonhardt, S, Veltzke-Schlieker, W, Adler, A, Schott, E, Hetzer, R, Schaffartzik, W, et al.. Trigger mechanisms of secondary sclerosing cholangitis in critically ill patients. Crit Care 2015;19:131. https://doi.org/10.1186/s13054-015-0861-5.Search in Google Scholar PubMed PubMed Central
113. Shlomo, E, Gadi, L, Daniel, S. Post-traumatic secondary biliary cirrhosis. Harefuah 2009;148:235–7. 77.Search in Google Scholar
114. Saluja, S, White, RI. Hereditary hemorrhagic telangiectasia of the liver: hyperperfusion with relative ischemia--poverty amidst plenty. Radiology 2004;230:25–7. https://doi.org/10.1148/radiol.2293031482.Search in Google Scholar PubMed
115. Vlachou, PA, Colak, E, Koculym, A, Kirpalani, A, Kim, TK, Hirschfield, GM, et al.. Improvement of ischemic cholangiopathy in three patients with hereditary hemorrhagic telangiectasia following treatment with bevacizumab. J Hepatol 2013;59:186–9. https://doi.org/10.1016/j.jhep.2013.02.006.Search in Google Scholar PubMed
116. Goritsas, CP, Repanti, M, Papadaki, E, Lazarou, N, Andonopoulos, AP. Intrahepatic bile duct injury and nodular regenerative hyperplasia of the liver in a patient with polyarteritis nodosa. J Hepatol 1997;26:727–30. https://doi.org/10.1016/s0168-8278(97)80441-2.Search in Google Scholar PubMed
117. Choi, HL, Sung, RH, Kang, MH, Jeon, HJ, Yun, HY, Jang, LC, et al.. Polyarteritis nodosa presented as a dilatation of the intrahepatic bile duct. Ann Surg Treat Res 2014;87:273–5. https://doi.org/10.4174/astr.2014.87.5.273.Search in Google Scholar PubMed PubMed Central
118. Saiura, A, Umekita, N, Inoue, S, Maeshiro, T, Miyamoto, S, Matsui, Y, et al.. Benign biliary stricture associated with atherosclerosis. Hepato-Gastroenterology 2001;48:81–2.Search in Google Scholar
119. Issa, H, Al-Salem, AH. Hepatobiliary manifestations of sickle cell anemia. Gastroenterol Res 2010;3:1–8. https://doi.org/10.4021/gr2010.01.1332.Search in Google Scholar PubMed PubMed Central
120. Viola, S, Meyer, M, Fabre, M, Tounian, P, Goddon, R, Dechelotte, P, et al.. Ischemic necrosis of bile ducts complicating Schönlein-Henoch purpura. Gastroenterology 1999;117:211–4. https://doi.org/10.1016/s0016-5085(99)70569-x.Search in Google Scholar PubMed
121. Ebert, EC, Hagspiel, KD. Gastrointestinal and hepatic manifestations of systemic lupus erythematosus. J Clin Gastroenterol 2011;45:436–41. https://doi.org/10.1097/mcg.0b013e31820f81b8.Search in Google Scholar
122. Kaushik, S, Federle, MP, Schur, PH, Krishnan, M, Silverman, SG, Ros, PR. Abdominal thrombotic and ischemic manifestations of the antiphospholipid antibody syndrome: CT findings in 42 patients. Radiology 2001;218:768–71. https://doi.org/10.1148/radiology.218.3.r01fe43768.Search in Google Scholar PubMed
123. Durand, JM, Sahel, J, Barthet, M, Alliot, P, Argeme, M, Soubeyrand, J. Cholangitis and paroxysmal nocturnal hemoglobinuria. Dig Dis Sci 1997;42:279–80. https://doi.org/10.1023/a:1018897331227.10.1023/A:1018897331227Search in Google Scholar
124. Brunner, SM, Junger, H, Ruemmele, P, Schnitzbauer, AA, Doenecke, A, Kirchner, GI, et al.. Bile duct damage after cold storage of deceased donor livers predicts biliary complications after liver transplantation. J Hepatol 2013;58:1133–9. https://doi.org/10.1016/j.jhep.2012.12.022.Search in Google Scholar PubMed
125. Karimian, N, Westerkamp, AC, Porte, RJ. Biliary complications after orthotopic liver transplantation. Curr Opin Organ Transplant 2014;19:209–16. https://doi.org/10.1097/mot.0000000000000082.Search in Google Scholar
126. Karimian, N, Weeder, PD, Bomfati, F, Gouw, AS, Porte, RJ. Preservation injury of the distal extrahepatic bile duct of donor livers is representative for injury of the intrahepatic bile ducts. J Hepatol 2015;63:284–7. https://doi.org/10.1016/j.jhep.2015.03.015.Search in Google Scholar PubMed
127. de Jong, IEM, Matton, APM, van Praagh, JB, van Haaften, WT, Wiersema-Buist, J, van Wijk, LA, et al.. Peribiliary glands are key in regeneration of the human biliary epithelium after severe bile duct injury. Hepatology 2019;69:1719–34. https://doi.org/10.1002/hep.30365.Search in Google Scholar PubMed PubMed Central
128. Rieder, F, Fiocchi, C, Rogler, G. Mechanisms, management, and treatment of fibrosis in patients with inflammatory bowel diseases. Gastroenterology 2017;152:340–50.e6. https://doi.org/10.1053/j.gastro.2016.09.047.Search in Google Scholar PubMed PubMed Central
129. Nho, RS, Ballinger, MN, Rojas, MM, Ghadiali, SN, Horowitz, JC. Biomechanical force and cellular stiffness in lung fibrosis. Am J Pathol 2022;192:750–61. https://doi.org/10.1016/j.ajpath.2022.02.001.Search in Google Scholar PubMed PubMed Central
130. Plotnikov, SV, Pasapera, AM, Sabass, B, Waterman, CM. Force fluctuations within focal adhesions mediate ECM-rigidity sensing to guide directed cell migration. Cell 2012;151:1513–27. https://doi.org/10.1016/j.cell.2012.11.034.Search in Google Scholar PubMed PubMed Central
131. Frangogiannis, NG. Cardiac fibrosis. Cardiovasc Res 2021;117:1450–88. https://doi.org/10.1093/cvr/cvaa324.Search in Google Scholar PubMed PubMed Central
132. Koudstaal, T, Funke-Chambour, M, Kreuter, M, Molyneaux, PL, Wijsenbeek, MS. Pulmonary fibrosis: from pathogenesis to clinical decision-making. Trends Mol Med 2023;29:1076–87. https://doi.org/10.1016/j.molmed.2023.08.010.Search in Google Scholar PubMed
133. Humphreys, BD. Mechanisms of renal fibrosis. Annu Rev Physiol 2018;80:309–26. https://doi.org/10.1146/annurev-physiol-022516-034227.Search in Google Scholar PubMed
134. Djiambou-Nganjeu, H. Relationship between portal HTN and cirrhosis as a cause for diabetes. J Transl Int Med 2019;7:79–83. https://doi.org/10.2478/jtim-2019-0009.Search in Google Scholar PubMed PubMed Central
135. Bell, SC, Mall, MA, Gutierrez, H, Macek, M, Madge, S, Davies, JC, et al.. The future of cystic fibrosis care: a global perspective. Lancet Respir Med 2020;8:65–124. https://doi.org/10.1016/s2213-2600(19)30337-6.Search in Google Scholar PubMed PubMed Central
136. Zhang, Y, Wu, Y, Shen, W, Wang, B, Yuan, X. Crosstalk between NK cells and hepatic stellate cells in liver fibrosis (Review). Mol Med Rep 2022;25. https://doi.org/10.3892/mmr.2022.12724.Search in Google Scholar PubMed PubMed Central
137. Sava, P, Ramanathan, A, Dobronyi, A, Peng, X, Sun, H, Ledesma-Mendoza, A, et al.. Human pericytes adopt myofibroblast properties in the microenvironment of the IPF lung. JCI Insight 2017;2:e96352. https://doi.org/10.1172/jci.insight.96352.Search in Google Scholar PubMed PubMed Central
138. Ober, EA, Lemaigre, FP. Development of the liver: insights into organ and tissue morphogenesis. J Hepatol 2018;68:1049–62. https://doi.org/10.1016/j.jhep.2018.01.005.Search in Google Scholar PubMed
139. Banales, JM, Huebert, RC, Karlsen, T, Strazzabosco, M, LaRusso, NF, Gores, GJ. Cholangiocyte pathobiology. Nat Rev Gastroenterol Hepatol 2019;16:269–81. https://doi.org/10.1038/s41575-019-0125-y.Search in Google Scholar PubMed PubMed Central
140. Tchorz, JS, Kinter, J, Müller, M, Tornillo, L, Heim, MH, Bettler, B. Notch2 signaling promotes biliary epithelial cell fate specification and tubulogenesis during bile duct development in mice. Hepatology 2009;50:871–9. https://doi.org/10.1002/hep.23048.Search in Google Scholar PubMed
141. Decaens, T, Godard, C, de Reyniès, A, Rickman, DS, Tronche, F, Couty, JP, et al.. Stabilization of beta-catenin affects mouse embryonic liver growth and hepatoblast fate. Hepatology 2008;47:247–58. https://doi.org/10.1002/hep.21952.Search in Google Scholar PubMed
142. Zhang, Y, Beachy, PA. Cellular and molecular mechanisms of Hedgehog signalling. Nat Rev Mol Cell Biol 2023;24:668–87. https://doi.org/10.1038/s41580-023-00591-1.Search in Google Scholar PubMed
143. Clotman, F, Jacquemin, P, Plumb-Rudewiez, N, Pierreux, CE, Van der Smissen, P, Dietz, HC, et al.. Control of liver cell fate decision by a gradient of TGF beta signaling modulated by Onecut transcription factors. Genes Dev 2005;19:1849–54. https://doi.org/10.1101/gad.340305.Search in Google Scholar PubMed PubMed Central
144. Yanai, M, Tatsumi, N, Hasunuma, N, Katsu, K, Endo, F, Yokouchi, Y. FGF signaling segregates biliary cell-lineage from chick hepatoblasts cooperatively with BMP4 and ECM components in vitro. Dev Dynam 2008;237:1268–83. https://doi.org/10.1002/dvdy.21520.Search in Google Scholar PubMed
145. Lemaigre, FP. Molecular mechanisms of biliary development. Prog Mol Biol Transl Sci 2010;97:103–26. https://doi.org/10.1016/b978-0-12-385233-5.00004-0.Search in Google Scholar PubMed
146. Li, L, Krantz, ID, Deng, Y, Genin, A, Banta, AB, Collins, CC, et al.. Alagille syndrome is caused by mutations in human Jagged1, which encodes a ligand for Notch1. Nat Genet 1997;16:243–51. https://doi.org/10.1038/ng0797-243.Search in Google Scholar PubMed
147. Merino-Azpitarte, M, Lozano, E, Perugorria, MJ, Esparza-Baquer, A, Erice, O, Santos-Laso, Á, et al.. SOX17 regulates cholangiocyte differentiation and acts as a tumor suppressor in cholangiocarcinoma. J Hepatol 2017;67:72–83. https://doi.org/10.1016/j.jhep.2017.02.017.Search in Google Scholar PubMed PubMed Central
148. Adams, JM, Huppert, KA, Castro, EC, Lopez, MF, Niknejad, N, Subramanian, S, et al.. Sox9 is a modifier of the liver disease severity in a mouse model of Alagille syndrome. Hepatology 2020;71:1331–49. https://doi.org/10.1002/hep.30912.Search in Google Scholar PubMed PubMed Central
149. Masyuk, AI, Masyuk, TV, Splinter, PL, Huang, BQ, Stroope, AJ, LaRusso, NF. Cholangiocyte cilia detect changes in luminal fluid flow and transmit them into intracellular Ca2+ and cAMP signaling. Gastroenterology 2006;131:911–20. https://doi.org/10.1053/j.gastro.2006.07.003.Search in Google Scholar PubMed PubMed Central
150. Overi, D, Carpino, G, Cardinale, V, Franchitto, A, Safarikia, S, Onori, P, et al.. Contribution of resident stem cells to liver and biliary tree regeneration in human diseases. Int J Mol Sci 2018;19. https://doi.org/10.3390/ijms19102917.Search in Google Scholar PubMed PubMed Central
151. Stanger, BZ. Cellular homeostasis and repair in the mammalian liver. Annu Rev Physiol 2015;77:179–200. https://doi.org/10.1146/annurev-physiol-021113-170255.Search in Google Scholar PubMed PubMed Central
152. Zong, Y, Stanger, BZ. Molecular mechanisms of liver and bile duct development. Wiley Interdiscip Rev Dev Biol 2012;1:643–55. https://doi.org/10.1002/wdev.47.Search in Google Scholar PubMed
153. Itoh, T, Miyajima, A. Liver regeneration by stem/progenitor cells. Hepatology 2014;59:1617–26. https://doi.org/10.1002/hep.26753.Search in Google Scholar PubMed
154. Choi, TY, Ninov, N, Stainier, DY, Shin, D. Extensive conversion of hepatic biliary epithelial cells to hepatocytes after near total loss of hepatocytes in zebrafish. Gastroenterology 2014;146:776–88. https://doi.org/10.1053/j.gastro.2013.10.019.Search in Google Scholar PubMed PubMed Central
155. He, J, Lu, H, Zou, Q, Luo, L. Regeneration of liver after extreme hepatocyte loss occurs mainly via biliary transdifferentiation in zebrafish. Gastroenterology 2014;146:789–800.e8. https://doi.org/10.1053/j.gastro.2013.11.045.Search in Google Scholar PubMed
156. Schaub, JR, Huppert, KA, Kurial, SNT, Hsu, BY, Cast, AE, Donnelly, B, et al.. De novo formation of the biliary system by TGFβ-mediated hepatocyte transdifferentiation. Nature 2018;557:247–51. https://doi.org/10.1038/s41586-018-0075-5.Search in Google Scholar PubMed PubMed Central
157. Masyuk, AI, Huang, BQ, Radtke, BN, Gajdos, GB, Splinter, PL, Masyuk, TV, et al.. Ciliary subcellular localization of TGR5 determines the cholangiocyte functional response to bile acid signaling. Am J Physiol Gastrointest Liver Physiol 2013;304:G1013–24. https://doi.org/10.1152/ajpgi.00383.2012.Search in Google Scholar PubMed PubMed Central
158. Masyuk, AI, Gradilone, SA, Banales, JM, Huang, BQ, Masyuk, TV, Lee, SO, et al.. Cholangiocyte primary cilia are chemosensory organelles that detect biliary nucleotides via P2Y12 purinergic receptors. Am J Physiol Gastrointest Liver Physiol 2008;295:G725–34. https://doi.org/10.1152/ajpgi.90265.2008.Search in Google Scholar PubMed PubMed Central
159. Masyuk, AI, Huang, BQ, Ward, CJ, Gradilone, SA, Banales, JM, Masyuk, TV, et al.. Biliary exosomes influence cholangiocyte regulatory mechanisms and proliferation through interaction with primary cilia. Am J Physiol Gastrointest Liver Physiol 2010;299:G990–9. https://doi.org/10.1152/ajpgi.00093.2010.Search in Google Scholar PubMed PubMed Central
160. Gradilone, SA, Masyuk, AI, Splinter, PL, Banales, JM, Huang, BQ, Tietz, PS, et al.. Cholangiocyte cilia express TRPV4 and detect changes in luminal tonicity inducing bicarbonate secretion. Proc Natl Acad Sci U S A 2007;104:19138–43. https://doi.org/10.1073/pnas.0705964104.Search in Google Scholar PubMed PubMed Central
161. Chen, XM, O’Hara, SP, Nelson, JB, Splinter, PL, Small, AJ, Tietz, PS, et al.. Multiple TLRs are expressed in human cholangiocytes and mediate host epithelial defense responses to Cryptosporidium parvum via activation of NF-kappaB. J Immunol 2005;175:7447–56. https://doi.org/10.4049/jimmunol.175.11.7447.Search in Google Scholar PubMed
162. Harada, K, Ohira, S, Isse, K, Ozaki, S, Zen, Y, Sato, Y, et al.. Lipopolysaccharide activates nuclear factor-kappaB through toll-like receptors and related molecules in cultured biliary epithelial cells. Lab Invest 2003;83:1657–67. https://doi.org/10.1097/01.lab.0000097190.56734.fe.Search in Google Scholar PubMed
163. Auth, MK, Keitzer, RA, Scholz, M, Blaheta, RA, Hottenrott, EC, Herrmann, G, et al.. Establishment and immunological characterization of cultured human gallbladder epithelial cells. Hepatology 1993;18:546–55. https://doi.org/10.1002/hep.1840180311.Search in Google Scholar
164. Godfrey, DI, Uldrich, AP, McCluskey, J, Rossjohn, J, Moody, DB. The burgeoning family of unconventional T cells. Nat Immunol 2015;16:1114–23. https://doi.org/10.1038/ni.3298.Search in Google Scholar PubMed
165. Schrumpf, E, Tan, C, Karlsen, TH, Sponheim, J, Björkström, NK, Sundnes, O, et al.. The biliary epithelium presents antigens to and activates natural killer T cells. Hepatology 2015;62:1249–59. https://doi.org/10.1002/hep.27840.Search in Google Scholar PubMed PubMed Central
166. Desmet, VJ. Histopathology of cholestasis. Verh Dtsch Ges Pathol 1995;79:233–40.Search in Google Scholar
167. Fabris, L, Strazzabosco, M. Epithelial-mesenchymal interactions in biliary diseases. Semin Liver Dis 2011;31:11–32. https://doi.org/10.1055/s-0031-1272832.Search in Google Scholar PubMed PubMed Central
168. Brivio, S, Cadamuro, M, Fabris, L, Strazzabosco, M. Epithelial-to-Mesenchymal transition and cancer invasiveness: what can we learn from cholangiocarcinoma? J Clin Med 2015;4:2028–41. https://doi.org/10.3390/jcm4121958.Search in Google Scholar PubMed PubMed Central
169. Fabris, L, Cadamuro, M, Guido, M, Spirli, C, Fiorotto, R, Colledan, M, et al.. Analysis of liver repair mechanisms in Alagille syndrome and biliary atresia reveals a role for notch signaling. Am J Pathol 2007;171:641–53. https://doi.org/10.2353/ajpath.2007.070073.Search in Google Scholar PubMed PubMed Central
170. Morell, CM, Fiorotto, R, Meroni, M, Raizner, A, Torsello, B, Cadamuro, M, et al.. Notch signaling and progenitor/ductular reaction in steatohepatitis. PLoS One 2017;12:e0187384. https://doi.org/10.1371/journal.pone.0187384.Search in Google Scholar PubMed PubMed Central
171. Geisler, F, Strazzabosco, M. Emerging roles of Notch signaling in liver disease. Hepatology 2015;61:382–92. https://doi.org/10.1002/hep.27268.Search in Google Scholar PubMed PubMed Central
172. Panciera, T, Azzolin, L, Cordenonsi, M, Piccolo, S. Mechanobiology of YAP and TAZ in physiology and disease. Nat Rev Mol Cell Biol 2017;18:758–70. https://doi.org/10.1038/nrm.2017.87.Search in Google Scholar PubMed PubMed Central
173. Francis, H, Glaser, S, Ueno, Y, Lesage, G, Marucci, L, Benedetti, A, et al.. cAMP stimulates the secretory and proliferative capacity of the rat intrahepatic biliary epithelium through changes in the PKA/Src/MEK/ERK1/2 pathway. J Hepatol 2004;41:528–37. https://doi.org/10.1016/j.jhep.2004.06.009.Search in Google Scholar PubMed
174. Roskams, TA, Libbrecht, L, Desmet, VJ. Progenitor cells in diseased human liver. Semin Liver Dis 2003;23:385–96. https://doi.org/10.1055/s-2004-815564.Search in Google Scholar PubMed
175. Sasaki, M, Ikeda, H, Yamaguchi, J, Miyakoshi, M, Sato, Y, Nakanuma, Y. Bile ductular cells undergoing cellular senescence increase in chronic liver diseases along with fibrous progression. Am J Clin Pathol 2010;133:212–23. https://doi.org/10.1309/ajcpwmx47treywzg.Search in Google Scholar PubMed
176. Moncsek, A, Al-Suraih, MS, Trussoni, CE, O’Hara, SP, Splinter, PL, Zuber, C, et al.. Targeting senescent cholangiocytes and activated fibroblasts with B-cell lymphoma-extra large inhibitors ameliorates fibrosis in multidrug resistance 2 gene knockout (Mdr2(-/-) ) mice. Hepatology 2018;67:247–59. https://doi.org/10.1002/hep.29464.Search in Google Scholar PubMed PubMed Central
177. Joshi, N, Kopec, AK, O’Brien, KM, Towery, KL, Cline-Fedewa, H, Williams, KJ, et al.. Coagulation-driven platelet activation reduces cholestatic liver injury and fibrosis in mice. J Thromb Haemostasis 2015;13:57–71. https://doi.org/10.1111/jth.12770.Search in Google Scholar PubMed PubMed Central
178. Sato, K, Hall, C, Glaser, S, Francis, H, Meng, F, Alpini, G. Pathogenesis of kupffer cells in cholestatic liver injury. Am J Pathol 2016;186:2238–47. https://doi.org/10.1016/j.ajpath.2016.06.003.Search in Google Scholar PubMed PubMed Central
179. Laskin, DL, Sunil, VR, Gardner, CR, Laskin, JD. Macrophages and tissue injury: agents of defense or destruction? Annu Rev Pharmacol Toxicol 2011;51:267–88. https://doi.org/10.1146/annurev.pharmtox.010909.105812.Search in Google Scholar PubMed PubMed Central
180. Sica, A, Mantovani, A. Macrophage plasticity and polarization: in vivo veritas. J Clin Invest 2012;122:787–95. https://doi.org/10.1172/jci59643.Search in Google Scholar
181. Johnson, C, Han, Y, Hughart, N, McCarra, J, Alpini, G, Meng, F. Interleukin-6 and its receptor, key players in hepatobiliary inflammation and cancer. Transl Gastrointest Cancer 2012;1:58–70. https://doi.org/10.3978/j.issn.2224-4778.2011.11.02.Search in Google Scholar PubMed PubMed Central
182. Ponomarev, ED, Veremeyko, T, Barteneva, N, Krichevsky, AM, Weiner, HL. MicroRNA-124 promotes microglia quiescence and suppresses EAE by deactivating macrophages via the C/EBP-α-PU.1 pathway. Nat Med 2011;17:64–70. https://doi.org/10.1038/nm.2266.Search in Google Scholar PubMed PubMed Central
183. Zhuang, G, Meng, C, Guo, X, Cheruku, PS, Shi, L, Xu, H, et al.. A novel regulator of macrophage activation: miR-223 in obesity-associated adipose tissue inflammation. Circulation 2012;125:2892–903. https://doi.org/10.1161/circulationaha.111.087817.Search in Google Scholar PubMed
184. Martinez-Nunez, RT, Louafi, F, Sanchez-Elsner, T. The interleukin 13 (IL-13) pathway in human macrophages is modulated by microRNA-155 via direct targeting of interleukin 13 receptor alpha1 (IL13Ralpha1). J Biol Chem 2011;286:1786–94. https://doi.org/10.1074/jbc.m110.169367.Search in Google Scholar PubMed PubMed Central
185. Ma, PF, Gao, CC, Yi, J, Zhao, JL, Liang, SQ, Zhao, Y, et al.. Cytotherapy with M1-polarized macrophages ameliorates liver fibrosis by modulating immune microenvironment in mice. J Hepatol 2017;67:770–9. https://doi.org/10.1016/j.jhep.2017.05.022.Search in Google Scholar PubMed
186. Wan, J, Benkdane, M, Teixeira-Clerc, F, Bonnafous, S, Louvet, A, Lafdil, F, et al.. M2 Kupffer cells promote M1 Kupffer cell apoptosis: a protective mechanism against alcoholic and nonalcoholic fatty liver disease. Hepatology 2014;59:130–42. https://doi.org/10.1002/hep.26607.Search in Google Scholar PubMed
187. De Muynck, K, Vanderborght, B, De Ponti, FF, Gijbels, E, Van Welden, S, Guilliams, M, et al.. Kupffer cells contested as early drivers in the pathogenesis of primary sclerosing cholangitis. Am J Pathol 2023;193:366–79. https://doi.org/10.1016/j.ajpath.2022.12.008.Search in Google Scholar PubMed
188. Golbar, HM, Izawa, T, Bondoc, A, Wijesundera, KK, Tennakoon, AH, Kuwamura, M, et al.. Attenuation of alpha-naphthylisothiocyanate (ANIT)-induced biliary fibrosis by depletion of hepatic macrophages in rats. Exp Toxicol Pathol 2017;69:221–30. https://doi.org/10.1016/j.etp.2017.01.005.Search in Google Scholar PubMed
189. Li, X, Liu, R, Wang, Y, Zhu, W, Zhao, D, Wang, X, et al.. Cholangiocyte-derived exosomal lncRNA H19 promotes macrophage activation and hepatic inflammation under cholestatic conditions. Cells 2020;9:190. https://doi.org/10.3390/cells9010190.Search in Google Scholar PubMed PubMed Central
190. Tian, X, Wang, Y, Lu, Y, Wang, W, Du, J, Chen, S, et al.. Conditional depletion of macrophages ameliorates cholestatic liver injury and fibrosis via lncRNA-H19. Cell Death Dis 2021;12:646. https://doi.org/10.1038/s41419-021-03931-1.Search in Google Scholar PubMed PubMed Central
191. Zhou, T, Meadows, V, Kundu, D, Kyritsi, K, Owen, T, Ceci, L, et al.. Mast cells selectively target large cholangiocytes during biliary injury via H2HR-mediated cAMP/pERK1/2 signaling. Hepatol Commun 2022;6:2715–31. https://doi.org/10.1002/hep4.2026.Search in Google Scholar PubMed PubMed Central
192. Hargrove, L, Kennedy, L, Demieville, J, Jones, H, Meng, F, DeMorrow, S, et al.. Bile duct ligation-induced biliary hyperplasia, hepatic injury, and fibrosis are reduced in mast cell-deficient Kit(W-sh) mice. Hepatology 2017;65:1991–2004. https://doi.org/10.1002/hep.29079.Search in Google Scholar PubMed PubMed Central
193. Yang, M, Vanderwert, E, Kimchi, ET, Staveley-O’Carroll, KF, Li, G. The important roles of natural killer cells in liver fibrosis. Biomedicines 2023;11. https://doi.org/10.3390/biomedicines11051391.Search in Google Scholar PubMed PubMed Central
194. Duman, DG, Zibandeh, N, Ugurlu, MU, Celikel, C, Akkoc, T, Banzragch, M, et al.. Mesenchymal stem cells suppress hepatic fibrosis accompanied by expanded intrahepatic natural killer cells in rat fibrosis model. Mol Biol Rep 2019;46:2997–3008. https://doi.org/10.1007/s11033-019-04736-4.Search in Google Scholar PubMed
195. Santodomingo-Garzon, T, Swain, MG. Role of NKT cells in autoimmune liver disease. Autoimmun Rev 2011;10:793–800. https://doi.org/10.1016/j.autrev.2011.06.003.Search in Google Scholar PubMed
196. Godfrey, DI, Hammond, KJ, Poulton, LD, Smyth, MJ, Baxter, AG. NKT cells: facts, functions and fallacies. Immunol Today 2000;21:573–83. https://doi.org/10.1016/s0167-5699(00)01735-7.Search in Google Scholar PubMed
197. Tasneem, AA, Luck, NH. Autoimmune hepatitis: clinical characteristics and predictors of biochemical response to treatment. J Transl Int Med 2020;8:106–11. https://doi.org/10.2478/jtim-2020-0016.Search in Google Scholar PubMed PubMed Central
198. Aso-Ishimoto, Y, Yamagiwa, S, Ichida, T, Miyakawa, R, Tomiyama, C, Sato, Y, et al.. Increased activated natural killer T cells in the liver of patients with advanced stage primary biliary cirrhosis. Biomed Res 2014;35:161–9. https://doi.org/10.2220/biomedres.35.161.Search in Google Scholar PubMed
199. Berntsen, NL, Fosby, B, Tan, C, Reims, HM, Ogaard, J, Jiang, X, et al.. Natural killer T cells mediate inflammation in the bile ducts. Mucosal Immunol 2018;11:1582–90. https://doi.org/10.1038/s41385-018-0066-8.Search in Google Scholar PubMed PubMed Central
200. Jia, H, Chen, J, Zhang, X, Bi, K, Zhou, H, Liu, T, et al.. IL-17A produced by invariant natural killer T cells and CD3(+) CD56(+) αGalcer-CD1d tetramer(-) T cells promote liver fibrosis in patients with primary biliary cholangitis. J Leukoc Biol 2022;112:1079–87. https://doi.org/10.1002/jlb.2a0622-586rrrr.Search in Google Scholar PubMed
201. Tsuneyama, K, Baba, H, Morimoto, Y, Tsunematsu, T, Ogawa, H. Primary biliary cholangitis: its pathological characteristics and immunopathological mechanisms. J Med Invest 2017;64:7–13. https://doi.org/10.2152/jmi.64.7.Search in Google Scholar PubMed
202. Sakamoto, N, Muraji, T, Ohtani, H, Masumoto, K. The accumulation of regulatory T cells in the hepatic hilar lymph nodes in biliary atresia. Surg Today 2017;47:1282–6. https://doi.org/10.1007/s00595-017-1502-1.Search in Google Scholar PubMed
203. Behairy, BE, Ehsan, N, Anwer, M, Allam, A, El-Henawy, I, Hameed, NA, et al.. Expression of intrahepatic CD3, CD4, and CD8 T cells in biliary atresia. Clin Exp Hepatol 2018;4:7–12. https://doi.org/10.5114/ceh.2017.71394.Search in Google Scholar PubMed PubMed Central
204. Wen, J, Zhou, Y, Wang, J, Chen, J, Yan, W, Wu, J, et al.. Interactions between Th1 cells and Tregs affect regulation of hepatic fibrosis in biliary atresia through the IFN-γ/STAT1 pathway. Cell Death Differ 2017;24:997–1006. https://doi.org/10.1038/cdd.2017.31.Search in Google Scholar PubMed PubMed Central
205. Zhang, BB, Yan, C, Fang, F, Du, Y, Ma, R, Li, XY, et al.. Increased hepatic Th2 and Treg subsets are associated with biliary fibrosis in different strains of mice caused by Clonorchis sinensis. PLoS One 2017;12:e0171005. https://doi.org/10.1371/journal.pone.0171005.Search in Google Scholar PubMed PubMed Central
206. Zepeda-Morales, AS, Del Toro-Arreola, S, García-Benavides, L, Bastidas-Ramírez, BE, Fafutis-Morris, M, Pereira-Suárez, AL, et al.. Liver fibrosis in bile duct-ligated rats correlates with increased hepatic IL-17 and TGF-β2 expression. Ann Hepatol 2016;15:418–26. https://doi.org/10.5604/16652681.1198820.Search in Google Scholar PubMed
207. Nosalski, R, Siedlinski, M, Denby, L, McGinnigle, E, Nowak, M, Cat, AND, et al.. T-Cell-Derived miRNA-214 mediates perivascular fibrosis in hypertension. Circ Res 2020;126:988–1003. https://doi.org/10.1161/circresaha.119.315428.Search in Google Scholar PubMed PubMed Central
208. Taylor, AE, Carey, AN, Kudira, R, Lages, CS, Shi, T, Lam, S, et al.. Interleukin 2 promotes hepatic regulatory T cell responses and protects from biliary fibrosis in murine sclerosing cholangitis. Hepatology 2018;68:1905–21. https://doi.org/10.1002/hep.30061.Search in Google Scholar PubMed PubMed Central
209. Perugino, CA, Stone, JH. IgG4-related disease: an update on pathophysiology and implications for clinical care. Nat Rev Rheumatol 2020;16:702–14. https://doi.org/10.1038/s41584-020-0500-7.Search in Google Scholar PubMed
210. Cargill, T, Culver, EL. The role of B cells and B cell therapies in immune-mediated liver diseases. Front Immunol 2021;12:661196. https://doi.org/10.3389/fimmu.2021.661196.Search in Google Scholar PubMed PubMed Central
211. Della-Torre, E, Rigamonti, E, Perugino, C, Baghai-Sain, S, Sun, N, Kaneko, N, et al.. B lymphocytes directly contribute to tissue fibrosis in patients with IgG(4)-related disease. J Allergy Clin Immunol 2020;145:968–81.e14. https://doi.org/10.1016/j.jaci.2019.07.004.Search in Google Scholar PubMed PubMed Central
212. Maillette de Buy Wenniger, LJ, Doorenspleet, ME, Klarenbeek, PL, Verheij, J, Baas, F, Elferink, RP, et al.. Immunoglobulin G4+ clones identified by next-generation sequencing dominate the B cell receptor repertoire in immunoglobulin G4 associated cholangitis. Hepatology 2013;57:2390–8. https://doi.org/10.1002/hep.26232.Search in Google Scholar PubMed
213. Kuwahara, T, Hara, K, Mizuno, N, Haba, S, Okuno, N, Fukui, T, et al.. Current status of artificial intelligence analysis for the treatment of pancreaticobiliary diseases using endoscopic ultrasonography and endoscopic retrograde cholangiopancreatography. DEN Open 2024;4:e267. https://doi.org/10.1002/deo2.267.Search in Google Scholar PubMed PubMed Central
214. Muhl, L, Genové, G, Leptidis, S, Liu, J, He, L, Mocci, G, et al.. Single-cell analysis uncovers fibroblast heterogeneity and criteria for fibroblast and mural cell identification and discrimination. Nat Commun 2020;11:3953. https://doi.org/10.1038/s41467-020-17740-1.Search in Google Scholar PubMed PubMed Central
215. Hynes, RO, Naba, A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harbor Perspect Biol 2012;4:a004903. https://doi.org/10.1101/cshperspect.a004903.Search in Google Scholar PubMed PubMed Central
216. Huang, X, Yang, N, Fiore, VF, Barker, TH, Sun, Y, Morris, SW, et al.. Matrix stiffness-induced myofibroblast differentiation is mediated by intrinsic mechanotransduction. Am J Respir Cell Mol Biol 2012;47:340–8. https://doi.org/10.1165/rcmb.2012-0050oc.Search in Google Scholar
217. Junker, JP, Sommar, P, Skog, M, Johnson, H, Kratz, G. Adipogenic, chondrogenic and osteogenic differentiation of clonally derived human dermal fibroblasts. Cells Tissues Organs 2010;191:105–18. https://doi.org/10.1159/000232157.Search in Google Scholar PubMed
218. Rivera-Gonzalez, GC, Shook, BA, Andrae, J, Holtrup, B, Bollag, K, Betsholtz, C, et al.. Skin adipocyte stem cell self-renewal is regulated by a PDGFA/AKT-Signaling Axis. Cell Stem Cell 2016;19:738–51. https://doi.org/10.1016/j.stem.2016.09.002.Search in Google Scholar PubMed PubMed Central
219. Buechler, MB, Pradhan, RN, Krishnamurty, AT, Cox, C, Calviello, AK, Wang, AW, et al.. Cross-tissue organization of the fibroblast lineage. Nature 2021;593:575–9. https://doi.org/10.1038/s41586-021-03549-5.Search in Google Scholar PubMed
220. Pakshir, P, Alizadehgiashi, M, Wong, B, Coelho, NM, Chen, X, Gong, Z, et al.. Dynamic fibroblast contractions attract remote macrophages in fibrillar collagen matrix. Nat Commun 2019;10:1850. https://doi.org/10.1038/s41467-019-09709-6.Search in Google Scholar PubMed PubMed Central
221. Hinz, B. The myofibroblast: paradigm for a mechanically active cell. J Biomech 2010;43:146–55. https://doi.org/10.1016/j.jbiomech.2009.09.020.Search in Google Scholar PubMed
222. Tschumperlin, DJ. Fibroblasts and the ground they walk on. Physiology 2013;28:380–90. https://doi.org/10.1152/physiol.00024.2013.Search in Google Scholar PubMed PubMed Central
223. Ferrer, RA, Saalbach, A, Grünwedel, M, Lohmann, N, Forstreuter, I, Saupe, S, et al.. Dermal fibroblasts promote alternative macrophage activation improving impaired wound healing. J Invest Dermatol 2017;137:941–50. https://doi.org/10.1016/j.jid.2016.11.035.Search in Google Scholar PubMed
224. Uezumi, A, Fukada, S, Yamamoto, N, Takeda, S, Tsuchida, K. Mesenchymal progenitors distinct from satellite cells contribute to ectopic fat cell formation in skeletal muscle. Nat Cell Biol 2010;12:143–52. https://doi.org/10.1038/ncb2014.Search in Google Scholar PubMed
225. Nakaya, M, Watari, K, Tajima, M, Nakaya, T, Matsuda, S, Ohara, H, et al.. Cardiac myofibroblast engulfment of dead cells facilitates recovery after myocardial infarction. J Clin Invest 2017;127:383–401. https://doi.org/10.1172/jci83822.Search in Google Scholar
226. Guerrero-Juarez, CF, Dedhia, PH, Jin, S, Ruiz-Vega, R, Ma, D, Liu, Y, et al.. Single-cell analysis reveals fibroblast heterogeneity and myeloid-derived adipocyte progenitors in murine skin wounds. Nat Commun 2019;10:650. https://doi.org/10.1038/s41467-018-08247-x.Search in Google Scholar PubMed PubMed Central
227. Opalenik, SR, Davidson, JM. Fibroblast differentiation of bone marrow-derived cells during wound repair. Faseb J 2005;19:1561–3. https://doi.org/10.1096/fj.04-2978fje.Search in Google Scholar PubMed
228. Sinha, M, Sen, CK, Singh, K, Das, A, Ghatak, S, Rhea, B, et al.. Direct conversion of injury-site myeloid cells to fibroblast-like cells of granulation tissue. Nat Commun 2018;9:936. https://doi.org/10.1038/s41467-018-03208-w.Search in Google Scholar PubMed PubMed Central
229. Kruglikov, IL, Scherer, PE. Adipocyte-myofibroblast transition as a possible pathophysiological step in androgenetic alopecia. Exp Dermatol 2017;26:522–3. https://doi.org/10.1111/exd.13379.Search in Google Scholar PubMed
230. Marangoni, RG, Korman, BD, Wei, J, Wood, TA, Graham, LV, Whitfield, ML, et al.. Myofibroblasts in murine cutaneous fibrosis originate from adiponectin-positive intradermal progenitors. Arthritis Rheumatol 2015;67:1062–73. https://doi.org/10.1002/art.38990.Search in Google Scholar PubMed PubMed Central
231. Shook, BA, Wasko, RR, Mano, O, Rutenberg-Schoenberg, M, Rudolph, MC, Zirak, B, et al.. Dermal adipocyte lipolysis and myofibroblast conversion are required for efficient skin repair. Cell Stem Cell 2020;26:880–95.e6. https://doi.org/10.1016/j.stem.2020.03.013.Search in Google Scholar PubMed PubMed Central
232. Hung, C, Linn, G, Chow, YH, Kobayashi, A, Mittelsteadt, K, Altemeier, WA, et al.. Role of lung pericytes and resident fibroblasts in the pathogenesis of pulmonary fibrosis. Am J Respir Crit Care Med 2013;188:820–30. https://doi.org/10.1164/rccm.201212-2297oc.Search in Google Scholar
233. Kuppe, C, Ibrahim, MM, Kranz, J, Zhang, X, Ziegler, S, Perales-Patón, J, et al.. Decoding myofibroblast origins in human kidney fibrosis. Nature 2021;589:281–6. https://doi.org/10.1038/s41586-020-2941-1.Search in Google Scholar PubMed PubMed Central
234. Lua, I, James, D, Wang, J, Wang, KS, Asahina, K. Mesodermal mesenchymal cells give rise to myofibroblasts, but not epithelial cells, in mouse liver injury. Hepatology 2014;60:311–22. https://doi.org/10.1002/hep.27035.Search in Google Scholar PubMed PubMed Central
235. Zagory, JA, Nguyen, MV, Dietz, W, Mavila, N, Haldeman, A, Grishin, A, et al.. Toll-like receptor 3 mediates PROMININ-1 expressing cell expansion in biliary atresia via Transforming Growth Factor-Beta. J Pediatr Surg 2016;51:917–22. https://doi.org/10.1016/j.jpedsurg.2016.02.054.Search in Google Scholar PubMed
236. Copple, BL. Hypoxia stimulates hepatocyte epithelial to mesenchymal transition by hypoxia-inducible factor and transforming growth factor-beta-dependent mechanisms. Liver Int 2010;30:669–82. https://doi.org/10.1111/j.1478-3231.2010.02205.x.Search in Google Scholar PubMed PubMed Central
237. Qiu, BF, Zhang, GQ, Xu, FM, Xu, Q, Xu, T. Effect of the transdifferentiation of BECs into myofibroblasts on the pathogenesis of secondary cholestatic hepatic fibrosis. Exp Ther Med 2019;17:2769–76. https://doi.org/10.3892/etm.2019.7234.Search in Google Scholar PubMed PubMed Central
238. Rygiel, KA, Robertson, H, Marshall, HL, Pekalski, M, Zhao, L, Booth, TA, et al.. Epithelial-mesenchymal transition contributes to portal tract fibrogenesis during human chronic liver disease. Lab Invest 2008;88:112–23. https://doi.org/10.1038/labinvest.3700704.Search in Google Scholar PubMed
239. Koyama, Y, Wang, P, Liang, S, Iwaisako, K, Liu, X, Xu, J, et al.. Mesothelin/mucin 16 signaling in activated portal fibroblasts regulates cholestatic liver fibrosis. J Clin Invest 2017;127:1254–70. https://doi.org/10.1172/jci88845.Search in Google Scholar
240. Dranoff, JA, Wells, RG. Portal fibroblasts: underappreciated mediators of biliary fibrosis. Hepatology 2010;51:1438–44. https://doi.org/10.1002/hep.23405.Search in Google Scholar PubMed PubMed Central
241. Wells, RG, Kruglov, E, Dranoff, JA. Autocrine release of TGF-beta by portal fibroblasts regulates cell growth. FEBS Lett 2004;559:107–10. https://doi.org/10.1016/s0014-5793(04)00037-7.Search in Google Scholar
242. El Mourabit, H, Loeuillard, E, Lemoinne, S, Cadoret, A, Housset, C. Culture model of rat portal myofibroblasts. Front Physiol 2016;7:120. https://doi.org/10.3389/fphys.2016.00120.Search in Google Scholar PubMed PubMed Central
243. Michelotti, GA, Tucker, A, Swiderska-Syn, M, Machado, MV, Choi, SS, Kruger, L, et al.. Pleiotrophin regulates the ductular reaction by controlling the migration of cells in liver progenitor niches. Gut 2016;65:683–92. https://doi.org/10.1136/gutjnl-2014-308176.Search in Google Scholar PubMed PubMed Central
244. Asawa, S, Saito, T, Satoh, A, Ohtake, K, Tsuchiya, T, Okada, H, et al.. Participation of bone marrow cells in biliary fibrosis after bile duct ligation. J Gastroenterol Hepatol 2007;22:2001–8. https://doi.org/10.1111/j.1440-1746.2006.04708.x.Search in Google Scholar PubMed
245. Su, J, Morgani, SM, David, CJ, Wang, Q, Er, EE, Huang, YH, et al.. TGF-β orchestrates fibrogenic and developmental EMTs via the RAS effector RREB1. Nature 2020;577:566–71. https://doi.org/10.1038/s41586-019-1897-5.Search in Google Scholar PubMed PubMed Central
246. Jung, SW, Kim, SM, Kim, A, Park, SH, Moon, JY, Lee, SH. Midbody plays an active role in fibroblast-myofibroblast transition by mediating TGF-β signaling. Faseb J 2022;36:e22272. https://doi.org/10.1096/fj.202101613r.Search in Google Scholar PubMed
247. Massagué, J, Sheppard, D. TGF-β signaling in health and disease. Cell 2023;186:4007–37. https://doi.org/10.1016/j.cell.2023.07.036.Search in Google Scholar PubMed PubMed Central
248. Xu, S, Mao, Y, Wu, J, Feng, J, Li, J, Wu, L, et al.. TGF-β/Smad and JAK/STAT pathways are involved in the anti-fibrotic effects of propylene glycol alginate sodium sulphate on hepatic fibrosis. J Cell Mol Med 2020;24:5224–37. https://doi.org/10.1111/jcmm.15175.Search in Google Scholar PubMed PubMed Central
249. Xiang, D, Zou, J, Zhu, X, Chen, X, Luo, J, Kong, L, et al.. Physalin D attenuates hepatic stellate cell activation and liver fibrosis by blocking TGF-β/Smad and YAP signaling. Phytomedicine 2020;78:153294. https://doi.org/10.1016/j.phymed.2020.153294.Search in Google Scholar PubMed
250. Dropmann, A, Dooley, S, Dewidar, B, Hammad, S, Dediulia, T, Werle, J, et al.. TGF-β2 silencing to target biliary-derived liver diseases. Gut 2020;69:1677–90. https://doi.org/10.1136/gutjnl-2019-319091.Search in Google Scholar PubMed PubMed Central
251. Wang, P, Lei, S, Wang, X, Xu, W, Hu, P, Chen, F, et al.. MicroRNA-134 deactivates hepatic stellate cells by targeting TGF-β activated kinase 1-binding protein 1. Biochem Cell Biol 2019;97:505–12. https://doi.org/10.1139/bcb-2018-0211.Search in Google Scholar PubMed
252. Tang, X, Xie, X, Wang, X, Wang, Y, Jiang, X, Jiang, H. The combination of piR-823 and eukaryotic initiation factor 3 B (EIF3B) activates hepatic stellate cells via upregulating TGF-β1 in liver fibrogenesis. Med Sci Monit 2018;24:9151–65. https://doi.org/10.12659/msm.914222.Search in Google Scholar
253. Qian, J, Jiao, Y, Wang, G, Liu, H, Cao, X, Yang, H. Mechanism of TGF-β1 inhibiting Kupffer cell immune responses in cholestatic cirrhosis. Exp Ther Med 2020;20:1541–9. https://doi.org/10.3892/etm.2020.8826.Search in Google Scholar PubMed PubMed Central
254. Li, S, Zhou, B, Xue, M, Zhu, J, Tong, G, Fan, J, et al.. Macrophage-specific FGF12 promotes liver fibrosis progression in mice. Hepatology 2023;77:816–33. https://doi.org/10.1002/hep.32640.Search in Google Scholar PubMed
255. O’Brien, A, Zhou, T, White, T, Medford, A, Chen, L, Kyritsi, K, et al.. FGF1 signaling modulates biliary injury and liver fibrosis in the Mdr2(-/-) mouse model of primary sclerosing cholangitis. Hepatol Commun 2022;6:1574–88. https://doi.org/10.1002/hep4.1909.Search in Google Scholar PubMed PubMed Central
256. Hui, Q, Jin, Z, Li, X, Liu, C, Wang, X. FGF family: from drug development to clinical application. Int J Mol Sci 2018;19:1875. https://doi.org/10.3390/ijms19071875.Search in Google Scholar PubMed PubMed Central
257. Chen, C, Li, X, Wang, L. Thymosinβ4 alleviates cholestatic liver fibrosis in mice through downregulating PDGF/PDGFR and TGFβ/Smad pathways. Dig Liver Dis 2020;52:324–30. https://doi.org/10.1016/j.dld.2019.08.014.Search in Google Scholar PubMed
258. Martin, IV, Borkham-Kamphorst, E, Zok, S, van Roeyen, CR, Eriksson, U, Boor, P, et al.. Platelet-derived growth factor (PDGF)-C neutralization reveals differential roles of PDGF receptors in liver and kidney fibrosis. Am J Pathol 2013;182:107–17. https://doi.org/10.1016/j.ajpath.2012.09.006.Search in Google Scholar PubMed
259. Borkham-Kamphorst, E, van Roeyen, CR, Ostendorf, T, Floege, J, Gressner, AM, Weiskirchen, R. Pro-fibrogenic potential of PDGF-D in liver fibrosis. J Hepatol 2007;46:1064–74. https://doi.org/10.1016/j.jhep.2007.01.029.Search in Google Scholar PubMed
260. Cai, Y, Huang, G, Ma, L, Dong, L, Chen, S, Shen, X, et al.. Smurf2, an E3 ubiquitin ligase, interacts with PDE4B and attenuates liver fibrosis through miR-132 mediated CTGF inhibition. Biochim Biophys Acta Mol Cell Res 2018;1865:297–308. https://doi.org/10.1016/j.bbamcr.2017.10.011.Search in Google Scholar PubMed
261. Rachfal, AW, Brigstock, DR. Connective tissue growth factor (CTGF/CCN2) in hepatic fibrosis. Hepatol Res 2003;26:1–9. https://doi.org/10.1016/s1386-6346(03)00115-3.Search in Google Scholar PubMed
262. Jia, S, Liu, X, Li, W, Xie, J, Yang, L, Li, L. Peroxisome proliferator-activated receptor gamma negatively regulates the differentiation of bone marrow-derived mesenchymal stem cells toward myofibroblasts in liver fibrogenesis. Cell Physiol Biochem 2015;37:2085–100. https://doi.org/10.1159/000438567.Search in Google Scholar PubMed
263. Lu, L, Wang, J, Lu, H, Zhang, G, Liu, Y, Wang, J, et al.. MicroRNA-130a and -130b enhance activation of hepatic stellate cells by suppressing PPARγ expression: a rat fibrosis model study. Biochem Biophys Res Commun 2015;465:387–93. https://doi.org/10.1016/j.bbrc.2015.08.012.Search in Google Scholar PubMed
264. Yang, L, Stimpson, SA, Chen, L, Wallace Harrington, W, Rockey, DC. Effectiveness of the PPARγ agonist, GW570, in liver fibrosis. Inflamm Res 2010;59:1061–71. https://doi.org/10.1007/s00011-010-0226-0.Search in Google Scholar PubMed
265. Tian, L, Wang, Y, Jang, YY. Wnt signaling in biliary development, proliferation, and fibrosis. Exp Biol Med 2022;247:360–7. https://doi.org/10.1177/15353702211061376.Search in Google Scholar PubMed PubMed Central
266. Nishio, T, Hu, R, Koyama, Y, Liang, S, Rosenthal, SB, Yamamoto, G, et al.. Activated hepatic stellate cells and portal fibroblasts contribute to cholestatic liver fibrosis in MDR2 knockout mice. J Hepatol 2019;71:573–85. https://doi.org/10.1016/j.jhep.2019.04.012.Search in Google Scholar PubMed
267. Loeuillard, E, El Mourabit, H, Lei, L, Lemoinne, S, Housset, C, Cadoret, A. Endoplasmic reticulum stress induces inverse regulations of major functions in portal myofibroblasts during liver fibrosis progression. Biochim Biophys Acta, Mol Basis Dis 2018;1864:3688–96. https://doi.org/10.1016/j.bbadis.2018.10.008.Search in Google Scholar PubMed
268. Lee, HS, Choi, J, Son, T, Wee, HJ, Bae, SJ, Seo, JH, et al.. Altered AKAP12 expression in portal fibroblasts and liver sinusoids mediates transition from hepatic fibrogenesis to fibrosis resolution. Exp Mol Med 2018;50:1–13. https://doi.org/10.1038/s12276-018-0074-5.Search in Google Scholar PubMed PubMed Central
269. Borkham-Kamphorst, E, Steffen, BT, van de Leur, E, Haas, U, Weiskirchen, R. Portal myofibroblasts are sensitive to CCN-mediated endoplasmic reticulum stress-related apoptosis with potential to attenuate biliary fibrogenesis. Cell Signal 2018;51:72–85. https://doi.org/10.1016/j.cellsig.2018.07.005.Search in Google Scholar PubMed
270. Borkham-Kamphorst, E, Schaffrath, C, Van de Leur, E, Haas, U, Tihaa, L, Meurer, SK, et al.. The anti-fibrotic effects of CCN1/CYR61 in primary portal myofibroblasts are mediated through induction of reactive oxygen species resulting in cellular senescence, apoptosis and attenuated TGF-β signaling. Biochim Biophys Acta 2014;1843:902–14. https://doi.org/10.1016/j.bbamcr.2014.01.023.Search in Google Scholar PubMed
271. Wilson, CL, Murphy, LB, Leslie, J, Kendrick, S, French, J, Fox, CR, et al.. Ubiquitin C-terminal hydrolase 1: a novel functional marker for liver myofibroblasts and a therapeutic target in chronic liver disease. J Hepatol 2015;63:1421–8. https://doi.org/10.1016/j.jhep.2015.07.034.Search in Google Scholar PubMed PubMed Central
272. Abbasi, S, Sinha, S, Labit, E, Rosin, NL, Yoon, G, Rahmani, W, et al.. Distinct regulatory programs control the latent regenerative potential of dermal fibroblasts during wound healing. Cell Stem Cell 2020;27:396–412.e6. https://doi.org/10.1016/j.stem.2020.07.008.Search in Google Scholar PubMed
273. Phan, QM, Fine, GM, Salz, L, Herrera, GG, Wildman, B, Driskell, IM, et al.. Lef1 expression in fibroblasts maintains developmental potential in adult skin to regenerate wounds. Elife 2020;9. https://doi.org/10.7554/elife.60066.Search in Google Scholar PubMed PubMed Central
274. Phan, QM, Sinha, S, Biernaskie, J, Driskell, RR. Single-cell transcriptomic analysis of small and large wounds reveals the distinct spatial organization of regenerative fibroblasts. Exp Dermatol 2021;30:92–101. https://doi.org/10.1111/exd.14244.Search in Google Scholar PubMed PubMed Central
275. Morrison, SJ, Spradling, AC. Stem cells and niches: mechanisms that promote stem cell maintenance throughout life. Cell 2008;132:598–611. https://doi.org/10.1016/j.cell.2008.01.038.Search in Google Scholar PubMed PubMed Central
276. Hinz, B, Phan, SH, Thannickal, VJ, Prunotto, M, Desmoulière, A, Varga, J, et al.. Recent developments in myofibroblast biology: paradigms for connective tissue remodeling. Am J Pathol 2012;180:1340–55. https://doi.org/10.1016/j.ajpath.2012.02.004.Search in Google Scholar PubMed PubMed Central
277. Demaria, M, Ohtani, N, Youssef, SA, Rodier, F, Toussaint, W, Mitchell, JR, et al.. An essential role for senescent cells in optimal wound healing through secretion of PDGF-AA. Dev Cell 2014;31:722–33. https://doi.org/10.1016/j.devcel.2014.11.012.Search in Google Scholar PubMed PubMed Central
278. Wilkinson, HN, Clowes, C, Banyard, KL, Matteuci, P, Mace, KA, Hardman, MJ. Elevated local senescence in diabetic wound healing is linked to pathological repair via CXCR2. J Invest Dermatol 2019;139:1171–81.e6. https://doi.org/10.1016/j.jid.2019.01.005.Search in Google Scholar PubMed
279. Plikus, MV, Guerrero-Juarez, CF, Ito, M, Li, YR, Dedhia, PH, Zheng, Y, et al.. Regeneration of fat cells from myofibroblasts during wound healing. Science 2017;355:748–52. https://doi.org/10.1126/science.aai8792.Search in Google Scholar PubMed PubMed Central
280. Hinz, B. Formation and function of the myofibroblast during tissue repair. J Invest Dermatol 2007;127:526–37. https://doi.org/10.1038/sj.jid.5700613.Search in Google Scholar PubMed
281. Sabino, J, Vieira-Silva, S, Machiels, K, Joossens, M, Falony, G, Ballet, V, et al.. Primary sclerosing cholangitis is characterised by intestinal dysbiosis independent from IBD. Gut 2016;65:1681–9. https://doi.org/10.1136/gutjnl-2015-311004.Search in Google Scholar PubMed PubMed Central
282. Kummen, M, Holm, K, Anmarkrud, JA, Nygård, S, Vesterhus, M, Høivik, ML, et al.. The gut microbial profile in patients with primary sclerosing cholangitis is distinct from patients with ulcerative colitis without biliary disease and healthy controls. Gut 2017;66:611–9. https://doi.org/10.1136/gutjnl-2015-310500.Search in Google Scholar PubMed
283. Bajer, L, Kverka, M, Kostovcik, M, Macinga, P, Dvorak, J, Stehlikova, Z, et al.. Distinct gut microbiota profiles in patients with primary sclerosing cholangitis and ulcerative colitis. World J Gastroenterol 2017;23:4548–58. https://doi.org/10.3748/wjg.v23.i25.4548.Search in Google Scholar PubMed PubMed Central
284. Iwasawa, K, Suda, W, Tsunoda, T, Oikawa-Kawamoto, M, Umetsu, S, Inui, A, et al.. Characterisation of the faecal microbiota in Japanese patients with paediatric-onset primary sclerosing cholangitis. Gut 2017;66:1344–6. https://doi.org/10.1136/gutjnl-2016-312533.Search in Google Scholar PubMed PubMed Central
285. Rühlemann, MC, Heinsen, FA, Zenouzi, R, Lieb, W, Franke, A, Schramm, C. Faecal microbiota profiles as diagnostic biomarkers in primary sclerosing cholangitis. Gut 2017;66:753–4. https://doi.org/10.1136/gutjnl-2016-312180.Search in Google Scholar PubMed
286. Torres, J, Palmela, C, Brito, H, Bao, X, Ruiqi, H, Moura-Santos, P, et al.. The gut microbiota, bile acids and their correlation in primary sclerosing cholangitis associated with inflammatory bowel disease. United European Gastroenterol J 2018;6:112–22. https://doi.org/10.1177/2050640617708953.Search in Google Scholar PubMed PubMed Central
287. Rühlemann, M, Liwinski, T, Heinsen, FA, Bang, C, Zenouzi, R, Kummen, M, et al.. Consistent alterations in faecal microbiomes of patients with primary sclerosing cholangitis independent of associated colitis. Aliment Pharmacol Ther 2019;50:580–9. https://doi.org/10.1111/apt.15375.Search in Google Scholar PubMed PubMed Central
288. Vieira-Silva, S, Sabino, J, Valles-Colomer, M, Falony, G, Kathagen, G, Caenepeel, C, et al.. Quantitative microbiome profiling disentangles inflammation- and bile duct obstruction-associated microbiota alterations across PSC/IBD diagnoses. Nat Microbiol 2019;4:1826–31. https://doi.org/10.1038/s41564-019-0483-9.Search in Google Scholar PubMed
289. Lemoinne, S, Kemgang, A, Ben Belkacem, K, Straube, M, Jegou, S, Corpechot, C, et al.. Fungi participate in the dysbiosis of gut microbiota in patients with primary sclerosing cholangitis. Gut 2020;69:92–102. https://doi.org/10.1136/gutjnl-2018-317791.Search in Google Scholar PubMed
290. Kummen, M, Thingholm, LB, Rühlemann, MC, Holm, K, Hansen, SH, Moitinho-Silva, L, et al.. Altered gut microbial metabolism of essential nutrients in primary sclerosing cholangitis. Gastroenterology 2021;160:1784–98.e0. https://doi.org/10.1053/j.gastro.2020.12.058.Search in Google Scholar PubMed PubMed Central
291. Denoth, L, Juillerat, P, Kremer, AE, Rogler, G, Scharl, M, Yilmaz, B, et al.. Modulation of the mucosa-associated microbiome linked to the PTPN2 risk gene in patients with primary sclerosing cholangitis and ulcerative colitis. Microorganisms 2021;9. https://doi.org/10.3390/microorganisms9081752.Search in Google Scholar PubMed PubMed Central
292. Liu, Q, Li, B, Li, Y, Wei, Y, Huang, B, Liang, J, et al.. Altered faecal microbiome and metabolome in IgG4-related sclerosing cholangitis and primary sclerosing cholangitis. Gut 2022;71:899–909. https://doi.org/10.1136/gutjnl-2020-323565.Search in Google Scholar PubMed
293. Lapidot, Y, Amir, A, Ben-Simon, S, Veitsman, E, Cohen-Ezra, O, Davidov, Y, et al.. Alterations of the salivary and fecal microbiome in patients with primary sclerosing cholangitis. Hepatol Int 2021;15:191–201. https://doi.org/10.1007/s12072-020-10089-z.Search in Google Scholar PubMed
294. Cortez, RV, Moreira, LN, Padilha, M, Bibas, MD, Toma, RK, Porta, G, et al.. Gut microbiome of children and adolescents with primary sclerosing cholangitis in association with ulcerative colitis. Front Immunol 2020;11:598152. https://doi.org/10.3389/fimmu.2020.598152.Search in Google Scholar PubMed PubMed Central
295. Miyabe, K, Chandrasekhara, V, Wongjarupong, N, Chen, J, Yang, L, Johnson, S, et al.. Potential role of inflammation-promoting biliary microbiome in primary sclerosing cholangitis and cholangiocarcinoma. Cancers 2022;14. https://doi.org/10.3390/cancers14092120.Search in Google Scholar PubMed PubMed Central
296. Liwinski, T, Zenouzi, R, John, C, Ehlken, H, Rühlemann, MC, Bang, C, et al.. Alterations of the bile microbiome in primary sclerosing cholangitis. Gut 2020;69:665–72. https://doi.org/10.1136/gutjnl-2019-318416.Search in Google Scholar PubMed
297. Furukawa, M, Moriya, K, Nakayama, J, Inoue, T, Momoda, R, Kawaratani, H, et al.. Gut dysbiosis associated with clinical prognosis of patients with primary biliary cholangitis. Hepatol Res 2020;50:840–52. https://doi.org/10.1111/hepr.13509.Search in Google Scholar PubMed
298. Zhou, YJ, Ying, GX, Dong, SL, Xiang, B, Jin, QF. Gut microbial profile of treatment-naive patients with primary biliary cholangitis. Front Immunol 2023;14:1126117. https://doi.org/10.3389/fimmu.2023.1126117.Search in Google Scholar PubMed PubMed Central
299. Tang, R, Wei, Y, Li, Y, Chen, W, Chen, H, Wang, Q, et al.. Gut microbial profile is altered in primary biliary cholangitis and partially restored after UDCA therapy. Gut 2018;67:534–41. https://doi.org/10.1136/gutjnl-2016-313332.Search in Google Scholar PubMed
300. Chen, W, Wei, Y, Xiong, A, Li, Y, Guan, H, Wang, Q, et al.. Comprehensive analysis of serum and fecal bile acid profiles and interaction with gut microbiota in primary biliary cholangitis. Clin Rev Allergy Immunol 2020;58:25–38. https://doi.org/10.1007/s12016-019-08731-2.Search in Google Scholar PubMed
301. Lammert, C, Shin, A, Xu, H, Hemmerich, C, O’Connell, TM, Chalasani, N. Short-chain fatty acid and fecal microbiota profiles are linked to fibrosis in primary biliary cholangitis. FEMS Microbiol Lett 2021;368:fnab038. https://doi.org/10.1093/femsle/fnab038.Search in Google Scholar PubMed PubMed Central
302. Abe, K, Takahashi, A, Fujita, M, Imaizumi, H, Hayashi, M, Okai, K, et al.. Dysbiosis of oral microbiota and its association with salivary immunological biomarkers in autoimmune liver disease. PLoS One 2018;13:e0198757. https://doi.org/10.1371/journal.pone.0198757.Search in Google Scholar PubMed PubMed Central
303. Lv, LX, Fang, DQ, Shi, D, Chen, DY, Yan, R, Zhu, YX, et al.. Alterations and correlations of the gut microbiome, metabolism and immunity in patients with primary biliary cirrhosis. Environ Microbiol 2016;18:2272–86. https://doi.org/10.1111/1462-2920.13401.Search in Google Scholar PubMed
304. Song, W, Sun, LY, Zhu, ZJ, Wei, L, Qu, W, Zeng, ZG, et al.. Association of gut microbiota and metabolites with disease progression in children with biliary atresia. Front Immunol 2021;12:698900. https://doi.org/10.3389/fimmu.2021.698900.Search in Google Scholar PubMed PubMed Central
305. Wang, J, Qian, T, Jiang, J, Yang, Y, Shen, Z, Huang, Y, et al.. Gut microbial profile in biliary atresia: a case-control study. J Gastroenterol Hepatol 2020;35:334–42. https://doi.org/10.1111/jgh.14777.Search in Google Scholar PubMed
306. Blesl, A, Jüngst, C, Lammert, F, Fauler, G, Rainer, F, Leber, B, et al.. Secondary sclerosing cholangitis in critically ill patients alters the gut-liver Axis: a case control study. Nutrients 2020;12:2728. https://doi.org/10.3390/nu12092728.Search in Google Scholar PubMed PubMed Central
307. Sakamoto, M, Tanaka, Y, Benno, Y, Ohkuma, M. Parabacteroides faecis sp. nov., isolated from human faeces. Int J Syst Evol Microbiol 2015;65:1342–6. https://doi.org/10.1099/ijs.0.000109.Search in Google Scholar PubMed
308. Narushima, S, Itoha, K, Miyamoto, Y, Park, SH, Nagata, K, Kuruma, K, et al.. Deoxycholic acid formation in gnotobiotic mice associated with human intestinal bacteria. Lipids 2006;41:835–43. https://doi.org/10.1007/s11745-006-5038-1.Search in Google Scholar PubMed
309. Zhao, L, Yang, R, Cheng, L, Wang, M, Jiang, Y, Wang, S. LPS-induced epithelial-mesenchymal transition of intrahepatic biliary epithelial cells. J Surg Res 2011;171:819–25. https://doi.org/10.1016/j.jss.2010.04.059.Search in Google Scholar PubMed
310. Fuchs, CD, Paumgartner, G, Mlitz, V, Kunczer, V, Halilbasic, E, Leditznig, N, et al.. Colesevelam attenuates cholestatic liver and bile duct injury in Mdr2(-/-) mice by modulating composition, signalling and excretion of faecal bile acids. Gut 2018;67:1683–91. https://doi.org/10.1136/gutjnl-2017-314553.Search in Google Scholar PubMed PubMed Central
311. Rupp, C, Friedrich, K, Folseraas, T, Wannhoff, A, Bode, KA, Weiss, KH, et al.. Fut2 genotype is a risk factor for dominant stenosis and biliary candida infections in primary sclerosing cholangitis. Aliment Pharmacol Ther 2014;39:873–82. https://doi.org/10.1111/apt.12663.Search in Google Scholar PubMed
312. Treeriya, R, Ho, PN, Titapun, A, Klanrit, P, Suksawat, M, Kulthawatsiri, T, et al.. (1)H NMR fecal metabolic phenotyping of periductal fibrosis- and cholangiocarcinoma-specific metabotypes defining perturbation in gut microbial-host co-metabolism. PeerJ 2023;11:e15386. https://doi.org/10.7717/peerj.15386.Search in Google Scholar PubMed PubMed Central
313. Liu, N, Li, D, Liu, D, Liu, Y, Lei, J. FOSL2 participates in renal fibrosis via SGK1-mediated epithelial-mesenchymal transition of proximal tubular epithelial cells. J Transl Int Med 2023;11:294–308. https://doi.org/10.2478/jtim-2023-0105.Search in Google Scholar PubMed PubMed Central
314. Zhou, B, Lin, W, Long, Y, Yang, Y, Zhang, H, Wu, K, et al.. Notch signaling pathway: architecture, disease, and therapeutics. Signal Transduct Targeted Ther 2022;7:95. https://doi.org/10.1038/s41392-022-00934-y.Search in Google Scholar PubMed PubMed Central
315. Rim, EY, Clevers, H, Nusse, R. The Wnt pathway: from signaling mechanisms to synthetic modulators. Annu Rev Biochem 2022;91:571–98. https://doi.org/10.1146/annurev-biochem-040320-103615.Search in Google Scholar PubMed
316. Jing, J, Wu, Z, Wang, J, Luo, G, Lin, H, Fan, Y, et al.. Hedgehog signaling in tissue homeostasis, cancers, and targeted therapies. Signal Transduct Targeted Ther 2023;8:315. https://doi.org/10.1038/s41392-023-01559-5.Search in Google Scholar PubMed PubMed Central
317. Siebel, C, Lendahl, U. Notch signaling in development, tissue homeostasis, and disease. Physiol Rev 2017;97:1235–94. https://doi.org/10.1152/physrev.00005.2017.Search in Google Scholar PubMed
318. Ni, MM, Wang, YR, Wu, WW, Xia, CC, Zhang, YH, Xu, J, et al.. Novel Insights on Notch signaling pathways in liver fibrosis. Eur J Pharmacol 2018;826:66–74. https://doi.org/10.1016/j.ejphar.2018.02.051.Search in Google Scholar PubMed
319. Russell, JO, Monga, SP. Wnt/β-Catenin signaling in liver development, homeostasis, and pathobiology. Annu Rev Pathol 2018;13:351–78. https://doi.org/10.1146/annurev-pathol-020117-044010.Search in Google Scholar PubMed PubMed Central
320. McDaniell, R, Warthen, DM, Sanchez-Lara, PA, Pai, A, Krantz, ID, Piccoli, DA, et al.. NOTCH2 mutations cause Alagille syndrome, a heterogeneous disorder of the notch signaling pathway. Am J Hum Genet 2006;79:169–73. https://doi.org/10.1086/505332.Search in Google Scholar PubMed PubMed Central
321. Yu, J, Zhu, C, Wang, X, Kim, K, Bartolome, A, Dongiovanni, P, et al.. Hepatocyte TLR4 triggers inter-hepatocyte Jagged1/Notch signaling to determine NASH-induced fibrosis. Sci Transl Med 2021;13. https://doi.org/10.1126/scitranslmed.abe1692.Search in Google Scholar PubMed PubMed Central
322. Tao, H, Yang, JJ, Shi, KH, Li, J. Wnt signaling pathway in cardiac fibrosis: new insights and directions. Metabolism 2016;65:30–40. https://doi.org/10.1016/j.metabol.2015.10.013.Search in Google Scholar PubMed
323. Miao, CG, Yang, YY, He, X, Huang, C, Huang, Y, Zhang, L, et al.. Wnt signaling in liver fibrosis: progress, challenges and potential directions. Biochimie 2013;95:2326–35. https://doi.org/10.1016/j.biochi.2013.09.003.Search in Google Scholar PubMed
324. Liu, T, Gonzalez De Los Santos, F, Hirsch, M, Wu, Z, Phan, SH. Noncanonical Wnt signaling promotes myofibroblast differentiation in pulmonary fibrosis. Am J Respir Cell Mol Biol 2021;65:489–99. https://doi.org/10.1165/rcmb.2020-0499oc.Search in Google Scholar
325. Kumar, V, Mondal, G, Dutta, R, Mahato, RI. Co-delivery of small molecule hedgehog inhibitor and miRNA for treating liver fibrosis. Biomaterials 2016;76:144–56. https://doi.org/10.1016/j.biomaterials.2015.10.047.Search in Google Scholar PubMed
326. Tzavlaki, K, Moustakas, A. TGF-Β signaling. Biomolecules 2020;10:487. https://doi.org/10.3390/biom10030487.Search in Google Scholar PubMed PubMed Central
327. Hu, HH, Chen, DQ, Wang, YN, Feng, YL, Cao, G, Vaziri, ND, et al.. New insights into TGF-β/Smad signaling in tissue fibrosis. Chem Biol Interact 2018;292:76–83. https://doi.org/10.1016/j.cbi.2018.07.008.Search in Google Scholar PubMed
328. Zhang, J, Tian, XJ, Xing, J. Signal transduction pathways of EMT induced by TGF-β, SHH, and WNT and their crosstalks. J Clin Med 2016;5:41. https://doi.org/10.3390/jcm5040041.Search in Google Scholar PubMed PubMed Central
329. Ying, HZ, Chen, Q, Zhang, WY, Zhang, HH, Ma, Y, Zhang, SZ, et al.. PDGF signaling pathway in hepatic fibrosis pathogenesis and therapeutics (Review). Mol Med Rep 2017;16:7879–89. https://doi.org/10.3892/mmr.2017.7641.Search in Google Scholar PubMed PubMed Central
330. Kim, TH, Moon, JH, Savard, CE, Kuver, R, Lee, SP. Effects of lipopolysaccharide on platelet-derived growth factor isoform and receptor expression in cultured rat common bile duct fibroblasts and cholangiocytes. J Gastroenterol Hepatol 2009;24:1218–25. https://doi.org/10.1111/j.1440-1746.2008.05729.x.Search in Google Scholar PubMed
331. Maddaluno, L, Urwyler, C, Werner, S. Fibroblast growth factors: key players in regeneration and tissue repair. Development 2017;144:4047–60. https://doi.org/10.1242/dev.152587.Search in Google Scholar PubMed
332. Zhou, Y, Zhu, X, Cui, H, Shi, J, Yuan, G, Shi, S, et al.. The role of the VEGF family in coronary heart disease. Front Cardiovasc Med 2021;8:738325. https://doi.org/10.3389/fcvm.2021.738325.Search in Google Scholar PubMed PubMed Central
333. Ahmad, A, Nawaz, MI. Molecular mechanism of VEGF and its role in pathological angiogenesis. J Cell Biochem 2022;123:1938–65. https://doi.org/10.1002/jcb.30344.Search in Google Scholar PubMed
334. Zhang, J, Chu, M. Differential roles of VEGF: relevance to tissue fibrosis. J Cell Biochem 2019;120:10945–51. https://doi.org/10.1002/jcb.28489.Search in Google Scholar PubMed
335. Ihn, H. Pathogenesis of fibrosis: role of TGF-beta and CTGF. Curr Opin Rheumatol 2002;14:681–5. https://doi.org/10.1097/00002281-200211000-00009.Search in Google Scholar PubMed
336. Gressner, OA, Gressner, AM. Connective tissue growth factor: a fibrogenic master switch in fibrotic liver diseases. Liver Int 2008;28:1065–79. https://doi.org/10.1111/j.1478-3231.2008.01826.x.Search in Google Scholar PubMed
337. Philips, RL, Wang, Y, Cheon, H, Kanno, Y, Gadina, M, Sartorelli, V, et al.. The JAK-STAT pathway at 30: much learned, much more to do. Cell 2022;185:3857–76. https://doi.org/10.1016/j.cell.2022.09.023.Search in Google Scholar PubMed PubMed Central
338. Mair, M, Blaas, L, Österreicher, CH, Casanova, E, Eferl, R. JAK-STAT signaling in hepatic fibrosis. Front Biosci 2011;16:2794–811. https://doi.org/10.2741/3886.Search in Google Scholar PubMed
339. Vallée, A, Lecarpentier, Y, Guillevin, R, Vallée, JN. Interactions between TGF-β1, canonical WNT/β-catenin pathway and PPAR γ in radiation-induced fibrosis. Oncotarget 2017;8:90579–604. https://doi.org/10.18632/oncotarget.21234.Search in Google Scholar PubMed PubMed Central
340. Muzio, M, Polntarutti, N, Bosisio, D, Prahladan, MK, Mantovani, A. Toll like receptor family (TLT) and signalling pathway. Eur Cytokine Netw 2000;11:489–90.10.1042/bst028a489bSearch in Google Scholar
341. Wang, Y, Shi, K, Tu, J, Ke, C, Chen, N, Wang, B, et al.. Atractylenolide III ameliorates bile duct ligation-induced liver fibrosis by inhibiting the PI3K/AKT pathway and regulating glutamine metabolism. Molecules 2023;28:5504. https://doi.org/10.3390/molecules28145504.Search in Google Scholar PubMed PubMed Central
342. Hartmann, P, Haimerl, M, Mazagova, M, Brenner, DA, Schnabl, B. Toll-like receptor 2-mediated intestinal injury and enteric tumor necrosis factor receptor I contribute to liver fibrosis in mice. Gastroenterology 2012;143:1330–40.e1. https://doi.org/10.1053/j.gastro.2012.07.099.Search in Google Scholar PubMed PubMed Central
343. Wang, Y, Wang, X, Zhang, N, Yu, Y, Bao, P, Ma, Y, et al.. Extracellular vesicles of Clonorchis sinensis promote IL-6 and TNF-α secretion via the Toll-like receptor 9-mediated ERK pathway in biliary epithelial cells. Dev Comp Immunol 2023;139:104555. https://doi.org/10.1016/j.dci.2022.104555.Search in Google Scholar PubMed
344. Gäbele, E, Mühlbauer, M, Dorn, C, Weiss, TS, Froh, M, Schnabl, B, et al.. Role of TLR9 in hepatic stellate cells and experimental liver fibrosis. Biochem Biophys Res Commun 2008;376:271–6. https://doi.org/10.1016/j.bbrc.2008.08.096.Search in Google Scholar PubMed
345. Chinetti-Gbaguidi, G, Fruchart, JC, Staels, B. Role of the PPAR family of nuclear receptors in the regulation of metabolic and cardiovascular homeostasis: new approaches to therapy. Curr Opin Pharmacol 2005;5:177–83. https://doi.org/10.1016/j.coph.2004.11.004.Search in Google Scholar PubMed
346. Quintero, P, Arrese, M. Nuclear control of inflammation and fibrosis in nonalcoholic steatohepatitis: therapeutic potential of dual peroxisome proliferator-activated receptor alpha/delta agonism. Hepatology 2013;58:1881–4. https://doi.org/10.1002/hep.26582.Search in Google Scholar PubMed
347. Lu, Q, Guo, P, Guo, J, Ares, I, Lopez-Torres, B, Martínez-Larrañaga, MR, et al.. Targeting peroxisome proliferator-activated receptors: a new strategy for the treatment of cardiac fibrosis. Pharmacol Ther 2021;219:107702. https://doi.org/10.1016/j.pharmthera.2020.107702.Search in Google Scholar PubMed
348. Zambrano, S, Blanca, AJ, Ruiz-Armenta, MV, Miguel-Carrasco, JL, Arévalo, M, Mate, A, et al.. L-carnitine attenuates the development of kidney fibrosis in hypertensive rats by upregulating PPAR-γ. Am J Hypertens 2014;27:460–70. https://doi.org/10.1093/ajh/hpt268.Search in Google Scholar PubMed
349. Lakatos, HF, Thatcher, TH, Kottmann, RM, Garcia, TM, Phipps, RP, Sime, PJ. The role of PPARs in lung fibrosis. PPAR Res 2007;2007:71323. https://doi.org/10.1155/2007/71323.Search in Google Scholar PubMed PubMed Central
350. Russell, DW. The enzymes, regulation, and genetics of bile acid synthesis. Annu Rev Biochem 2003;72:137–74. https://doi.org/10.1146/annurev.biochem.72.121801.161712.Search in Google Scholar PubMed
351. Deng, F, Bae, YH. Bile acid transporter-mediated oral drug delivery. J Contr Release 2020;327:100–16. https://doi.org/10.1016/j.jconrel.2020.07.034.Search in Google Scholar PubMed PubMed Central
352. Aseem, SO, Hylemon, PB, Zhou, H. Bile acids and biliary fibrosis. Cells 2023;12. https://doi.org/10.3390/cells12050792.Search in Google Scholar PubMed PubMed Central
353. Sun, L, Cai, J, Gonzalez, FJ. The role of farnesoid X receptor in metabolic diseases, and gastrointestinal and liver cancer. Nat Rev Gastroenterol Hepatol 2021;18:335–47. https://doi.org/10.1038/s41575-020-00404-2.Search in Google Scholar PubMed
354. Wang, XX, Jiang, T, Shen, Y, Adorini, L, Pruzanski, M, Gonzalez, FJ, et al.. The farnesoid X receptor modulates renal lipid metabolism and diet-induced renal inflammation, fibrosis, and proteinuria. Am J Physiol Ren Physiol 2009;297:F1587–96. https://doi.org/10.1152/ajprenal.00404.2009.Search in Google Scholar PubMed PubMed Central
355. Feng, S, Xie, X, Li, J, Xu, X, Chen, C, Zou, G, et al.. Bile acids induce liver fibrosis through the NLRP3 inflammasome pathway and the mechanism of FXR inhibition of NLRP3 activation. Hepatol Int 2024;18:1040–52. https://doi.org/10.1007/s12072-023-10610-0.Search in Google Scholar PubMed PubMed Central
356. Schumacher, JD, Kong, B, Wu, J, Rizzolo, D, Armstrong, LE, Chow, MD, et al.. Direct and indirect effects of fibroblast growth factor (FGF) 15 and FGF19 on liver fibrosis development. Hepatology 2020;71:670–85. https://doi.org/10.1002/hep.30810.Search in Google Scholar PubMed PubMed Central
357. Xue, R, Su, L, Lai, S, Wang, Y, Zhao, D, Fan, J, et al.. Bile acid receptors and the gut-liver Axis in nonalcoholic fatty liver disease. Cells 2021;10:2806. https://doi.org/10.3390/cells10112806.Search in Google Scholar PubMed PubMed Central
358. Keitel, V, Ullmer, C, Häussinger, D. The membrane-bound bile acid receptor TGR5 (Gpbar-1) is localized in the primary cilium of cholangiocytes. Biol Chem 2010;391:785–9. https://doi.org/10.1515/bc.2010.077.Search in Google Scholar
359. Reich, M, Deutschmann, K, Sommerfeld, A, Klindt, C, Kluge, S, Kubitz, R, et al.. TGR5 is essential for bile acid-dependent cholangiocyte proliferation in vivo and in vitro. Gut 2016;65:487–501. https://doi.org/10.1136/gutjnl-2015-309458.Search in Google Scholar PubMed
360. Deutschmann, K, Reich, M, Klindt, C, Dröge, C, Spomer, L, Häussinger, D, et al.. Bile acid receptors in the biliary tree: TGR5 in physiology and disease. Biochim Biophys Acta, Mol Basis Dis 2018;1864:1319–25. https://doi.org/10.1016/j.bbadis.2017.08.021.Search in Google Scholar PubMed
361. Hov, JR, Keitel, V, Laerdahl, JK, Spomer, L, Ellinghaus, E, ElSharawy, A, et al.. Mutational characterization of the bile acid receptor TGR5 in primary sclerosing cholangitis. PLoS One 2010;5:e12403. https://doi.org/10.1371/journal.pone.0012403.Search in Google Scholar PubMed PubMed Central
362. Li, T, Holmstrom, SR, Kir, S, Umetani, M, Schmidt, DR, Kliewer, SA, et al.. The G protein-coupled bile acid receptor, TGR5, stimulates gallbladder filling. Mol Endocrinol 2011;25:1066–71. https://doi.org/10.1210/me.2010-0460.Search in Google Scholar PubMed PubMed Central
363. Baghdasaryan, A, Claudel, T, Gumhold, J, Silbert, D, Adorini, L, Roda, A, et al.. Dual farnesoid X receptor/TGR5 agonist INT-767 reduces liver injury in the Mdr2-/- (Abcb4-/-) mouse cholangiopathy model by promoting biliary HCO⁻₃ output. Hepatology 2011;54:1303–12. https://doi.org/10.1002/hep.24537.Search in Google Scholar PubMed PubMed Central
364. Koyama, Y, Brenner, DA. Liver inflammation and fibrosis. J Clin Invest 2017;127:55–64. https://doi.org/10.1172/jci88881.Search in Google Scholar
365. Wan, Y, Zhou, T, Slevin, E, Koyama, S, Li, X, Harrison, K, et al.. Liver-specific deletion of microRNA-34a alleviates ductular reaction and liver fibrosis during experimental cholestasis. Faseb J 2023;37:e22731. https://doi.org/10.1096/fj.202201453r.Search in Google Scholar PubMed
366. Bu, X, Ding, W, Guo, S, Wang, Y, Feng, J, Wang, P, et al.. Differential expression of microRNAs in bile duct obstruction-induced liver fibrosis and the identification of a novel liver fibrosis marker miR-1295b-3p. Ann Transl Med 2023;11:22. https://doi.org/10.21037/atm-22-6416.Search in Google Scholar PubMed PubMed Central
367. Zhou, Q, Rong, C, Gu, T, Li, H, Wu, L, Zhuansun, X, et al.. Mesenchymal stem cells improve liver fibrosis and protect hepatocytes by promoting microRNA-148a-5p-mediated inhibition of Notch signaling pathway. Stem Cell Res Ther 2022;13:354. https://doi.org/10.1186/s13287-022-03030-8.Search in Google Scholar PubMed PubMed Central
368. Yang, YL, Wang, FS, Lin, HY, Huang, YH. Exogenous therapeutics of microrna-29a attenuates development of hepatic fibrosis in cholestatic animal model through regulation of phosphoinositide 3-kinase p85 alpha. Int J Mol Sci 2020;21:3636. https://doi.org/10.3390/ijms21103636.Search in Google Scholar PubMed PubMed Central
369. Huang, YH, Kuo, HC, Yang, YL, Wang, FS. MicroRNA-29a is a key regulon that regulates BRD4 and mitigates liver fibrosis in mice by inhibiting hepatic stellate cell activation. Int J Med Sci 2019;16:212–20. https://doi.org/10.7150/ijms.29930.Search in Google Scholar PubMed PubMed Central
370. Huang, YH, Yang, YL, Huang, FC, Tiao, MM, Lin, YC, Tsai, MH, et al.. MicroRNA-29a mitigation of endoplasmic reticulum and autophagy aberrance counteracts in obstructive jaundice-induced fibrosis in mice. Exp Biol Med 2018;243:13–21. https://doi.org/10.1177/1535370217741500.Search in Google Scholar PubMed PubMed Central
371. Yang, YL, Wang, FS, Li, SC, Tiao, MM, Huang, YH. MicroRNA-29a alleviates bile duct ligation exacerbation of hepatic fibrosis in mice through epigenetic control of methyltransferases. Int J Mol Sci 2017;18:192. https://doi.org/10.3390/ijms18010192.Search in Google Scholar PubMed PubMed Central
372. Li, SC, Wang, FS, Yang, YL, Tiao, MM, Chuang, JH, Huang, YH. Microarray study of pathway analysis expression profile associated with MicroRNA-29a with regard to murine cholestatic liver injuries. Int J Mol Sci 2016;17:324. https://doi.org/10.3390/ijms17030324.Search in Google Scholar PubMed PubMed Central
373. Wang, Q, Wei, S, Zhou, H, Li, L, Zhou, S, Shi, C, et al.. MicroRNA-98 inhibits hepatic stellate cell activation and attenuates liver fibrosis by regulating HLF expression. Front Cell Dev Biol 2020;8:513. https://doi.org/10.3389/fcell.2020.00513.Search in Google Scholar PubMed PubMed Central
374. Santos, AA, Afonso, MB, Ramiro, RS, Pires, D, Pimentel, M, Castro, RE, et al.. Host miRNA-21 promotes liver dysfunction by targeting small intestinal Lactobacillus in mice. Gut Microb 2020;12:1–18. https://doi.org/10.1080/19490976.2020.1840766.Search in Google Scholar PubMed PubMed Central
375. Afonso, MB, Rodrigues, PM, Simão, AL, Gaspar, MM, Carvalho, T, Borralho, P, et al.. miRNA-21 ablation protects against liver injury and necroptosis in cholestasis. Cell Death Differ 2018;25:857–72. https://doi.org/10.1038/s41418-017-0019-x.Search in Google Scholar PubMed PubMed Central
376. Ning, ZW, Luo, XY, Wang, GZ, Li, Y, Pan, MX, Yang, RQ, et al.. MicroRNA-21 mediates angiotensin II-induced liver fibrosis by activating NLRP3 inflammasome/IL-1β Axis via targeting Smad7 and Spry1. Antioxid Redox Signal 2017;27:1–20. https://doi.org/10.1089/ars.2016.6669.Search in Google Scholar PubMed PubMed Central
377. Glaser, S, Meng, F, Han, Y, Onori, P, Chow, BK, Francis, H, et al.. Secretin stimulates biliary cell proliferation by regulating expression of microRNA 125b and microRNA let7a in mice. Gastroenterology 2014;146:1795–808.e12. https://doi.org/10.1053/j.gastro.2014.02.030.Search in Google Scholar PubMed PubMed Central
378. McDaniel, K, Wu, N, Zhou, T, Huang, L, Sato, K, Venter, J, et al.. Amelioration of ductular reaction by stem cell derived extracellular vesicles in MDR2 knockout mice via lethal-7 microRNA. Hepatology 2019;69:2562–78. https://doi.org/10.1002/hep.30542.Search in Google Scholar PubMed PubMed Central
379. Brandon-Warner, E, Benbow, JH, Swet, JH, Feilen, NA, Culberson, CR, McKillop, IH, et al.. Adeno-associated virus serotype 2 vector-mediated reintroduction of microRNA-19b attenuates hepatic fibrosis. Hum Gene Ther 2018;29:674–86. https://doi.org/10.1089/hum.2017.035.Search in Google Scholar PubMed PubMed Central
380. Venugopal, SK, Jiang, J, Kim, TH, Li, Y, Wang, SS, Torok, NJ, et al.. Liver fibrosis causes downregulation of miRNA-150 and miRNA-194 in hepatic stellate cells, and their overexpression causes decreased stellate cell activation. Am J Physiol Gastrointest Liver Physiol 2010;298:G101–6. https://doi.org/10.1152/ajpgi.00220.2009.Search in Google Scholar PubMed PubMed Central
381. Zhao, R, Dong, R, Yang, Y, Wang, Y, Ma, J, Wang, J, et al.. MicroRNA-155 modulates bile duct inflammation by targeting the suppressor of cytokine signaling 1 in biliary atresia. Pediatr Res 2017;82:1007–16. https://doi.org/10.1038/pr.2017.87.Search in Google Scholar PubMed
382. Zhang, R, Wang, M, Lu, H, Wang, J, Han, X, Liu, Z, et al.. A miR-340/SPP1 axis inhibits the activation and proliferation of hepatic stellate cells by inhibiting the TGF-β1/Smads pathway. Adv Clin Exp Med 2023;32:469–79. https://doi.org/10.17219/acem/154996.Search in Google Scholar PubMed
383. Navarro-Corcuera, A, Sehrawat, TS, Jalan-Sakrikar, N, Gibbons, HR, Pirius, NE, Khanal, S, et al.. Long non-coding RNA ACTA2-AS1 promotes ductular reaction by interacting with the p300/ELK1 complex. J Hepatol 2022;76:921–33. https://doi.org/10.1016/j.jhep.2021.12.014.Search in Google Scholar PubMed PubMed Central
384. Xiao, Y, Liu, R, Li, X, Gurley, EC, Hylemon, PB, Lu, Y, et al.. Long noncoding RNA H19 contributes to cholangiocyte proliferation and cholestatic liver fibrosis in biliary atresia. Hepatology 2019;70:1658–73. https://doi.org/10.1002/hep.30698.Search in Google Scholar PubMed PubMed Central
385. Liu, R, Li, X, Zhu, W, Wang, Y, Zhao, D, Wang, X, et al.. Cholangiocyte-derived exosomal long noncoding RNA H19 promotes hepatic stellate cell activation and cholestatic liver fibrosis. Hepatology 2019;70:1317–35. https://doi.org/10.1002/hep.30662.Search in Google Scholar PubMed PubMed Central
386. Peng, X, Yang, L, Liu, H, Pang, S, Chen, Y, Fu, J, et al.. Identification of circulating MicroRNAs in biliary atresia by next-generation sequencing. J Pediatr Gastroenterol Nutr 2016;63:518–23. https://doi.org/10.1097/mpg.0000000000001194.Search in Google Scholar
387. Yan, C, Shen, LP, Ma, R, Li, B, Li, XY, Hua, H, et al.. Characterization and identification of differentially expressed microRNAs during the process of the peribiliary fibrosis induced by Clonorchis sinensis. Infect Genet Evol 2016;43:321–8. https://doi.org/10.1016/j.meegid.2016.06.009.Search in Google Scholar PubMed
388. Li, X, Liu, R, Yang, J, Sun, L, Zhang, L, Jiang, Z, et al.. The role of long noncoding RNA H19 in gender disparity of cholestatic liver injury in multidrug resistance 2 gene knockout mice. Hepatology 2017;66:869–84. https://doi.org/10.1002/hep.29145.Search in Google Scholar PubMed PubMed Central
389. Rahimpour, S, Nasiri-Toosi, M, Khalili, H, Ebrahimi-Daryani, N, Nouri-Taromlou, MK, Azizi, Z. A triple blinded, randomized, placebo-controlled clinical trial to evaluate the efficacy and safety of oral vancomycin in primary sclerosing cholangitis: a pilot study. J Gastrointestin Liver Dis 2016;25:457–64. https://doi.org/10.15403/jgld.2014.1121.254.rah.Search in Google Scholar PubMed
390. Ali, AH, Damman, J, Shah, SB, Davies, Y, Hurwitz, M, Stephen, M, et al.. Open-label prospective therapeutic clinical trials: oral vancomycin in children and adults with primary sclerosing cholangitis. Scand J Gastroenterol 2020;55:941–50. https://doi.org/10.1080/00365521.2020.1787501.Search in Google Scholar PubMed
391. Abarbanel, DN, Seki, SM, Davies, Y, Marlen, N, Benavides, JA, Cox, K, et al.. Immunomodulatory effect of vancomycin on Treg in pediatric inflammatory bowel disease and primary sclerosing cholangitis. J Clin Immunol 2013;33:397–406. https://doi.org/10.1007/s10875-012-9801-1.Search in Google Scholar PubMed PubMed Central
392. Vleggaar, FP, Monkelbaan, JF, van Erpecum, KJ. Probiotics in primary sclerosing cholangitis: a randomized placebo-controlled crossover pilot study. Eur J Gastroenterol Hepatol 2008;20:688–92. https://doi.org/10.1097/meg.0b013e3282f5197e.Search in Google Scholar
393. Allegretti, JR, Kassam, Z, Carrellas, M, Mullish, BH, Marchesi, JR, Pechlivanis, A, et al.. Fecal microbiota transplantation in patients with primary sclerosing cholangitis: a pilot clinical trial. Am J Gastroenterol 2019;114:1071–9. https://doi.org/10.14309/ajg.0000000000000115.Search in Google Scholar PubMed
394. Schattenberg, JM, Pares, A, Kowdley, KV, Heneghan, MA, Caldwell, S, Pratt, D, et al.. A randomized placebo-controlled trial of elafibranor in patients with primary biliary cholangitis and incomplete response to UDCA. J Hepatol 2021;74:1344–54. https://doi.org/10.1016/j.jhep.2021.01.013.Search in Google Scholar PubMed
395. Bowlus, CL, Galambos, MR, Aspinall, RJ, Hirschfield, GM, Jones, DEJ, Dörffel, Y, et al.. A phase II, randomized, open-label, 52-week study of seladelpar in patients with primary biliary cholangitis. J Hepatol 2022;77:353–64. https://doi.org/10.1016/j.jhep.2022.02.033.Search in Google Scholar PubMed
396. Trauner, M, Chung, C, Sterling, K, Liu, X, Lu, X, Xu, J, et al.. PRIMIS: design of a pivotal, randomized, phase 3 study evaluating the safety and efficacy of the nonsteroidal farnesoid X receptor agonist cilofexor in noncirrhotic patients with primary sclerosing cholangitis. BMC Gastroenterol 2023;23:75. https://doi.org/10.1186/s12876-023-02653-2.Search in Google Scholar PubMed PubMed Central
397. Trauner, M, Bowlus, CL, Gulamhusein, A, Hameed, B, Caldwell, SH, Shiffman, ML, et al.. Safety and sustained efficacy of the farnesoid X receptor (FXR) agonist cilofexor over a 96-week open-label extension in patients with PSC. Clin Gastroenterol Hepatol 2023;21:1552–60.e2. https://doi.org/10.1016/j.cgh.2022.07.024.Search in Google Scholar PubMed
398. Lindor, KD, Kowdley, KV, Luketic, VA, Harrison, ME, McCashland, T, Befeler, AS, et al.. High-dose ursodeoxycholic acid for the treatment of primary sclerosing cholangitis. Hepatology 2009;50:808–14. https://doi.org/10.1002/hep.23082.Search in Google Scholar PubMed PubMed Central
399. Corpechot, C, Chazouillères, O, Rousseau, A, Le Gruyer, A, Habersetzer, F, Mathurin, P, et al.. A placebo-controlled trial of bezafibrate in primary biliary cholangitis. N Engl J Med 2018;378:2171–81. https://doi.org/10.1056/nejmoa1714519.Search in Google Scholar PubMed
400. Kowdley, KV, Vuppalanchi, R, Levy, C, Floreani, A, Andreone, P, LaRusso, NF, et al.. A randomized, placebo-controlled, phase II study of obeticholic acid for primary sclerosing cholangitis. J Hepatol 2020;73:94–101. https://doi.org/10.1016/j.jhep.2020.02.033.Search in Google Scholar PubMed PubMed Central
401. Fickert, P, Hirschfield, GM, Denk, G, Marschall, HU, Altorjay, I, Färkkilä, M, et al.. norUrsodeoxycholic acid improves cholestasis in primary sclerosing cholangitis. J Hepatol 2017;67:549–58. https://doi.org/10.1016/j.jhep.2017.05.009.Search in Google Scholar PubMed
402. Kowdley, KV, Forman, L, Eksteen, B, Gunn, N, Sundaram, V, Landis, C, et al.. A randomized, dose-finding, proof-of-concept study of berberine ursodeoxycholate in patients with primary sclerosing cholangitis. Am J Gastroenterol 2022;117:1805–15. https://doi.org/10.14309/ajg.0000000000001956.Search in Google Scholar PubMed
403. Schramm, C, Wedemeyer, H, Mason, A, Hirschfield, GM, Levy, C, Kowdley, KV, et al.. Farnesoid X receptor agonist tropifexor attenuates cholestasis in a randomised trial in patients with primary biliary cholangitis. JHEP Rep 2022;4:100544. https://doi.org/10.1016/j.jhepr.2022.100544.Search in Google Scholar PubMed PubMed Central
404. Sanyal, AJ, Ling, L, Beuers, U, DePaoli, AM, Lieu, HD, Harrison, SA, et al.. Potent suppression of hydrophobic bile acids by aldafermin, an FGF19 analogue, across metabolic and cholestatic liver diseases. JHEP Rep 2021;3:100255. https://doi.org/10.1016/j.jhepr.2021.100255.Search in Google Scholar PubMed PubMed Central
405. Loomes, KM, Squires, RH, Kelly, D, Rajwal, S, Soufi, N, Lachaux, A, et al.. Maralixibat for the treatment of PFIC: long-term, IBAT inhibition in an open-label, Phase 2 study. Hepatol Commun 2022;6:2379–90. https://doi.org/10.1002/hep4.1980.Search in Google Scholar PubMed PubMed Central
406. Tabibian, JH, Gossard, A, El-Youssef, M, Eaton, JE, Petz, J, Jorgensen, R, et al.. Prospective clinical trial of rifaximin therapy for patients with primary sclerosing cholangitis. Am J Ther 2017;24:e56–3. https://doi.org/10.1097/mjt.0000000000000102.Search in Google Scholar PubMed PubMed Central
407. Eksteen, B, Bowlus, CL, Montano-Loza, AJ, Lefebvre, E, Fischer, L, Vig, P, et al.. Efficacy and safety of cenicriviroc in patients with primary sclerosing cholangitis: PERSEUS study. Hepatol Commun 2021;5:478–90. https://doi.org/10.1002/hep4.1619.Search in Google Scholar PubMed PubMed Central
408. Muir, AJ, Levy, C, Janssen, HLA, Montano-Loza, AJ, Shiffman, ML, Caldwell, S, et al.. Simtuzumab for primary sclerosing cholangitis: phase 2 study results with insights on the natural history of the disease. Hepatology 2019;69:684–98. https://doi.org/10.1002/hep.30237.Search in Google Scholar PubMed
409. Arndtz, K, Chen, YY, Rowe, A, Homer, V, Kirkham, A, Douglas-Pugh, J, et al.. Efficacy and Mechanism Evaluation. Monoclonal antibody BTT1023 targeting vascular adhesion protein 1 for treating primary sclerosing cholangitis: BUTEO single-arm Phase II trial. Southampton (UK): NIHR Journals Library opyright © 2022 Arndtz et al; 2022.10.3310/ZPNF4670Search in Google Scholar PubMed
410. Kimura, M, Ogawa, E, Harada, K, Imamura, J, Saio, M, Ikura, Y, et al.. Feasibility, safety and tolerability of the CREB-binding protein/β-catenin inhibitor OP-724 in patients with advanced primary biliary cholangitis: an investigator-initiated, open-label, non-randomised, two-centre, phase 1 study. BMJ Open Gastroenterol 2022;9:e001001. https://doi.org/10.1136/bmjgast-2022-001001.Search in Google Scholar PubMed PubMed Central
411. Graffner, H, Gillberg, PG, Rikner, L, Marschall, HU. The ileal bile acid transporter inhibitor A4250 decreases serum bile acids by interrupting the enterohepatic circulation. Aliment Pharmacol Ther 2016;43:303–10. https://doi.org/10.1111/apt.13457.Search in Google Scholar PubMed
412. Baghdasaryan, A, Fuchs, CD, Österreicher, CH, Lemberger, UJ, Halilbasic, E, Påhlman, I, et al.. Inhibition of intestinal bile acid absorption improves cholestatic liver and bile duct injury in a mouse model of sclerosing cholangitis. J Hepatol 2016;64:674–81. https://doi.org/10.1016/j.jhep.2015.10.024.Search in Google Scholar PubMed
413. Al-Dury, S, Wahlström, A, Wahlin, S, Langedijk, J, Elferink, RO, Ståhlman, M, et al.. Pilot study with IBAT inhibitor A4250 for the treatment of cholestatic pruritus in primary biliary cholangitis. Sci Rep 2018;8:6658. https://doi.org/10.1038/s41598-018-25214-0.Search in Google Scholar PubMed PubMed Central
414. Lammers, WJ, Hirschfield, GM, Corpechot, C, Nevens, F, Lindor, KD, Janssen, HL, et al.. Development and validation of a scoring system to predict outcomes of patients with primary biliary cirrhosis receiving ursodeoxycholic acid therapy. Gastroenterology 2015;149:1804–12.e4. https://doi.org/10.1053/j.gastro.2015.07.061.Search in Google Scholar PubMed
415. Lindor, KD, Bowlus, CL, Boyer, J, Levy, C, Mayo, M. Primary biliary cholangitis: 2018 practice guidance from the American association for the study of liver diseases. Hepatology 2019;69:394–419. https://doi.org/10.1002/hep.30145.Search in Google Scholar PubMed
416. Alpini, G, Baiocchi, L, Glaser, S, Ueno, Y, Marzioni, M, Francis, H, et al.. Ursodeoxycholate and tauroursodeoxycholate inhibit cholangiocyte growth and secretion of BDL rats through activation of PKC alpha. Hepatology 2002;35:1041–52. https://doi.org/10.1053/jhep.2002.32712.Search in Google Scholar PubMed
417. Hatano, R, Kawaguchi, K, Togashi, F, Sugata, M, Masuda, S, Asano, S. Ursodeoxycholic acid ameliorates intrahepatic cholestasis independent of biliary bicarbonate secretion in vil2(kd/kd) mice. Biol Pharm Bull 2017;40:34–42. https://doi.org/10.1248/bpb.b16-00529.Search in Google Scholar PubMed
418. Nevens, F, Andreone, P, Mazzella, G, Strasser, SI, Bowlus, C, Invernizzi, P, et al.. A placebo-controlled trial of obeticholic acid in primary biliary cholangitis. N Engl J Med 2016;375:631–43. https://doi.org/10.1056/nejmoa1509840.Search in Google Scholar
419. Kowdley, KV, Luketic, V, Chapman, R, Hirschfield, GM, Poupon, R, Schramm, C, et al.. A randomized trial of obeticholic acid monotherapy in patients with primary biliary cholangitis. Hepatology 2018;67:1890–902. https://doi.org/10.1002/hep.29569.Search in Google Scholar PubMed PubMed Central
420. Wagner, M, Fickert, P, Zollner, G, Fuchsbichler, A, Silbert, D, Tsybrovskyy, O, et al.. Role of farnesoid X receptor in determining hepatic ABC transporter expression and liver injury in bile duct-ligated mice. Gastroenterology 2003;125:825–38. https://doi.org/10.1016/s0016-5085(03)01068-0.Search in Google Scholar PubMed
421. Rao, J, Yang, C, Yang, S, Lu, H, Hu, Y, Lu, L, et al.. Deficiency of TGR5 exacerbates immune-mediated cholestatic hepatic injury by stabilizing the β-catenin destruction complex. Int Immunol 2020;32:321–34. https://doi.org/10.1093/intimm/dxaa002.Search in Google Scholar PubMed PubMed Central
422. Wang, Y, Aoki, H, Yang, J, Peng, K, Liu, R, Li, X, et al.. The role of sphingosine 1-phosphate receptor 2 in bile-acid-induced cholangiocyte proliferation and cholestasis-induced liver injury in mice. Hepatology 2017;65:2005–18. https://doi.org/10.1002/hep.29076.Search in Google Scholar PubMed PubMed Central
423. Tanaka, A, Hirohara, J, Nakano, T, Matsumoto, K, Chazouillères, O, Takikawa, H, et al.. Association of bezafibrate with transplant-free survival in patients with primary biliary cholangitis. J Hepatol 2021;75:565–71. https://doi.org/10.1016/j.jhep.2021.04.010.Search in Google Scholar PubMed
424. Khakoo, NS, Sultan, S, Reynolds, JM, Levy, C. Efficacy and safety of bezafibrate alone or in combination with ursodeoxycholic acid in primary biliary cholangitis: systematic review and meta-analysis. Dig Dis Sci 2023;68:1559–73. https://doi.org/10.1007/s10620-022-07704-4.Search in Google Scholar PubMed
425. Grigorian, AY, Mardini, HE, Corpechot, C, Poupon, R, Levy, C. Fenofibrate is effective adjunctive therapy in the treatment of primary biliary cirrhosis: a meta-analysis. Clin Res Hepatol Gastroenterol 2015;39:296–306. https://doi.org/10.1016/j.clinre.2015.02.011.Search in Google Scholar PubMed
426. Mayo, MJ, Vierling, JM, Bowlus, CL, Neff, GW, Galambos, MR, Gordon, SC, et al.. Long-term safety and efficacy of seladelpar in patients with primary biliary cholangitis (PBC): 2-year results from a long-term study. Hepatology 2021;74:71A–3A.Search in Google Scholar
427. Fuchs, CD, Trauner, M. Role of bile acids and their receptors in gastrointestinal and hepatic pathophysiology. Nat Rev Gastroenterol Hepatol 2022;19:432–50. https://doi.org/10.1038/s41575-021-00566-7.Search in Google Scholar PubMed
428. Vuppalanchi, R, Caldwell, SH, Pyrsopoulos, N, deLemos, AS, Rossi, S, Levy, C, et al.. Proof-of-concept study to evaluate the safety and efficacy of saroglitazar in patients with primary biliary cholangitis. J Hepatol 2022;76:75–85. https://doi.org/10.1016/j.jhep.2021.08.025.Search in Google Scholar PubMed
429. Al-Najjar, AH, Ayob, AR, Awad, AS. Role of lactoferrin in treatment of bile duct ligation-induced hepatic fibrosis in rats: impact on inflammation and TGF-β1/smad2/α SMA signaling pathway. J Clin Exp Hepatol 2023;13:428–36. https://doi.org/10.1016/j.jceh.2022.12.014.Search in Google Scholar PubMed PubMed Central
430. ShamsEldeen, AM, Al-Ani, B, Ebrahim, HA, Rashed, L, Badr, AM, Attia, A, et al.. Resveratrol suppresses cholestasis-induced liver injury and fibrosis in rats associated with the inhibition of TGFβ1-Smad3-miR21 axis and profibrogenic and hepatic injury biomarkers. Clin Exp Pharmacol Physiol 2021;48:1402–11. https://doi.org/10.1111/1440-1681.13546.Search in Google Scholar PubMed
431. Ji, J, Yu, Q, Dai, W, Wu, L, Feng, J, Zheng, Y, et al.. Apigenin alleviates liver fibrosis by inhibiting hepatic stellate cell activation and autophagy via TGF-β1/Smad3 and p38/PPARα pathways. PPAR Res 2021;2021:6651839. https://doi.org/10.1155/2021/6651839.Search in Google Scholar PubMed PubMed Central
432. Chilvery, S, Bansod, S, Saifi, MA, Godugu, C. Piperlongumine attenuates bile duct ligation-induced liver fibrosis in mice via inhibition of TGF-β1/Smad and EMT pathways. Int Immunopharmacol 2020;88:106909. https://doi.org/10.1016/j.intimp.2020.106909.Search in Google Scholar PubMed
433. Liu, N, Feng, J, Lu, X, Yao, Z, Liu, Q, Lv, Y, et al.. Isorhamnetin inhibits liver fibrosis by reducing autophagy and inhibiting extracellular matrix formation via the TGF-β1/Smad3 and TGF-β1/p38 MAPK pathways. Mediat Inflamm 2019;2019:6175091. https://doi.org/10.1155/2019/6175091.Search in Google Scholar PubMed PubMed Central
434. Sharawy, MH, Abdel-Rahman, N, Megahed, N, El-Awady, MS. Paclitaxel alleviates liver fibrosis induced by bile duct ligation in rats: role of TGF-β1, IL-10 and c-Myc. Life Sci 2018;211:245–51. https://doi.org/10.1016/j.lfs.2018.09.037.Search in Google Scholar PubMed
435. Kong, R, Wang, N, Luo, H, Lu, J. Hesperetin mitigates bile duct ligation-induced liver fibrosis by inhibiting extracellular matrix and cell apoptosis via the TGF-β1/Smad pathway. Curr Mol Med 2018;18:15–24. https://doi.org/10.2174/1566524018666180608084947.Search in Google Scholar PubMed
436. Liu, H, Dong, F, Li, G, Niu, M, Zhang, C, Han, Y, et al.. Liuweiwuling tablets attenuate BDL-induced hepatic fibrosis via modulation of TGF-β/Smad and NF-κB signaling pathways. J Ethnopharmacol 2018;210:232–41. https://doi.org/10.1016/j.jep.2017.08.029.Search in Google Scholar PubMed
437. Zhou, L, Dong, X, Wang, L, Shan, L, Li, T, Xu, W, et al.. Casticin attenuates liver fibrosis and hepatic stellate cell activation by blocking TGF-β/Smad signaling pathway. Oncotarget 2017;8:56267–80. https://doi.org/10.18632/oncotarget.17453.Search in Google Scholar PubMed PubMed Central
438. Zhou, Y, Tong, X, Ren, S, Wang, X, Chen, J, Mu, Y, et al.. Synergistic anti-liver fibrosis actions of total astragalus saponins and glycyrrhizic acid via TGF-β1/Smads signaling pathway modulation. J Ethnopharmacol 2016;190:83–90. https://doi.org/10.1016/j.jep.2016.06.011.Search in Google Scholar PubMed
439. Li, J, Li, X, Xu, W, Wang, S, Hu, Z, Zhang, Q, et al.. Antifibrotic effects of luteolin on hepatic stellate cells and liver fibrosis by targeting AKT/mTOR/p70S6K and TGFβ/Smad signalling pathways. Liver Int 2015;35:1222–33. https://doi.org/10.1111/liv.12638.Search in Google Scholar PubMed
440. Aldaba-Muruato, LR, Moreno, MG, Hernández-Mercado, E, Shibayama, M, Muriel, P. Secondary biliary cirrhosis in the rat is prevented by decreasing NF-κB nuclear translocation and TGF-β expression using allopurinol, an inhibitor of xanthine oxidase. Can J Physiol Pharmacol 2012;90:1469–78. https://doi.org/10.1139/y2012-125.Search in Google Scholar PubMed
441. Kennedy, LL, Meng, F, Venter, JK, Zhou, T, Karstens, WA, Hargrove, LA, et al.. Knockout of microRNA-21 reduces biliary hyperplasia and liver fibrosis in cholestatic bile duct ligated mice. Lab Invest 2016;96:1256–67. https://doi.org/10.1038/labinvest.2016.112.Search in Google Scholar PubMed PubMed Central
442. Tanaka, A, Tazuma, S, Okazaki, K, Nakazawa, T, Inui, K, Chiba, T, et al.. Clinical features, response to treatment, and outcomes of IgG4-related sclerosing cholangitis. Clin Gastroenterol Hepatol 2017;15:920–6.e3. https://doi.org/10.1016/j.cgh.2016.12.038.Search in Google Scholar PubMed
443. Drazilova, S, Veseliny, E, Lenartova, PD, Drazilova, D, Gazda, J, Grgurevic, I, et al.. IgG4-Related sclerosing cholangitis: rarely diagnosed, but not a rare disease. Can J Gastroenterol Hepatol 2021;2021:1959832. https://doi.org/10.1155/2021/1959832.Search in Google Scholar PubMed PubMed Central
444. Kemp, W, Majeed, A, Mitchell, J, Majumdar, A, Tse, E, Skoien, R, et al.. Management, outcomes and survival of an Australian IgG4-SC cohort: the MOSAIC study. Liver Int 2021;41:2934–43. https://doi.org/10.1111/liv.15036.Search in Google Scholar PubMed
445. Ali, AH, Bi, Y, Machicado, JD, Garg, S, Lennon, RJ, Zhang, L, et al.. The long-term outcomes of patients with immunoglobulin G4-related sclerosing cholangitis: the Mayo Clinic experience. J Gastroenterol 2020;55:1087–97. https://doi.org/10.1007/s00535-020-01714-7.Search in Google Scholar PubMed
446. Lanzillotta, M, Della-Torre, E, Wallace, ZS, Stone, JH, Karadag, O, Fernández-Codina, A, et al.. Efficacy and safety of rituximab for IgG4-related pancreato-biliary disease: a systematic review and meta-analysis. Pancreatology 2021;21:1395–401. https://doi.org/10.1016/j.pan.2021.06.009.Search in Google Scholar PubMed
447. Betancur-Vásquez, L, Gonzalez-Hurtado, D, Arango-Isaza, D, Rojas-Villarraga, A, Hernandez-Parra, D, Carmona, S, et al.. IgG4-related disease: is rituximab the best therapeutic strategy for cases refractory to conventional therapy? Results of a systematic review. Reumatol Clínica 2020;16:195–202. https://doi.org/10.1016/j.reumae.2018.11.019.Search in Google Scholar
448. Rautiainen, H, Kärkkäinen, P, Karvonen, AL, Nurmi, H, Pikkarainen, P, Nuutinen, H, et al.. Budesonide combined with UDCA to improve liver histology in primary biliary cirrhosis: a three-year randomized trial. Hepatology 2005;41:747–52. https://doi.org/10.1002/hep.20646.Search in Google Scholar PubMed
449. Boberg, KM, Chapman, RW, Hirschfield, GM, Lohse, AW, Manns, MP, Schrumpf, E. Overlap syndromes: the International Autoimmune Hepatitis Group (IAIHG) position statement on a controversial issue. J Hepatol 2011;54:374–85. https://doi.org/10.1016/j.jhep.2010.09.002.Search in Google Scholar PubMed
450. Ishii, M, Iwai, M, Harada, Y, Morikawa, T, Okanoue, T, Kishikawa, T, et al.. A role of mast cells for hepatic fibrosis in primary sclerosing cholangitis. Hepatol Res 2005;31:127–31. https://doi.org/10.1016/j.hepres.2005.01.007.Search in Google Scholar PubMed
451. Kennedy, L, Hargrove, L, Demieville, J, Karstens, W, Jones, H, DeMorrow, S, et al.. Blocking H1/H2 histamine receptors inhibits damage/fibrosis in Mdr2(-/-) mice and human cholangiocarcinoma tumorigenesis. Hepatology 2018;68:1042–56. https://doi.org/10.1002/hep.29898.Search in Google Scholar PubMed PubMed Central
452. Wan, Y, Meng, F, Wu, N, Zhou, T, Venter, J, Francis, H, et al.. Substance P increases liver fibrosis by differential changes in senescence of cholangiocytes and hepatic stellate cells. Hepatology 2017;66:528–41. https://doi.org/10.1002/hep.29138.Search in Google Scholar PubMed PubMed Central
453. Glaser, S, Gaudio, E, Renzi, A, Mancinelli, R, Ueno, Y, Venter, J, et al.. Knockout of the neurokinin-1 receptor reduces cholangiocyte proliferation in bile duct-ligated mice. Am J Physiol Gastrointest Liver Physiol 2011;301:G297–305. https://doi.org/10.1152/ajpgi.00418.2010.Search in Google Scholar PubMed PubMed Central
454. Begum, R, Thota, S, Abdulkadir, A, Kaur, G, Bagam, P, Batra, S. NADPH oxidase family proteins: signaling dynamics to disease management. Cell Mol Immunol 2022;19:660–86. https://doi.org/10.1038/s41423-022-00858-1.Search in Google Scholar PubMed PubMed Central
455. Zhan, SS, Jiang, JX, Wu, J, Halsted, C, Friedman, SL, Zern, MA, et al.. Phagocytosis of apoptotic bodies by hepatic stellate cells induces NADPH oxidase and is associated with liver fibrosis in vivo. Hepatology 2006;43:435–43. https://doi.org/10.1002/hep.21093.Search in Google Scholar PubMed
456. Xue, C, Chu, Q, Li, L. Research progress on the role of probiotics in acute liver failure. J Transl Int Med 2022;10:83–5. https://doi.org/10.2478/jtim-2021-0052.Search in Google Scholar PubMed PubMed Central
457. Zhang, X, Luo, X, Tian, L, Yue, P, Li, M, Liu, K, et al.. The gut microbiome dysbiosis and regulation by fecal microbiota transplantation: umbrella review. Front Microbiol 2023;14:1286429. https://doi.org/10.3389/fmicb.2023.1286429.Search in Google Scholar PubMed PubMed Central
458. Tabibian, JH, Weeding, E, Jorgensen, RA, Petz, JL, Keach, JC, Talwalkar, JA, et al.. Randomised clinical trial: vancomycin or metronidazole in patients with primary sclerosing cholangitis - a pilot study. Aliment Pharmacol Ther 2013;37:604–12. https://doi.org/10.1111/apt.12232.Search in Google Scholar PubMed
459. Färkkilä, M, Karvonen, AL, Nurmi, H, Nuutinen, H, Taavitsainen, M, Pikkarainen, P, et al.. Metronidazole and ursodeoxycholic acid for primary sclerosing cholangitis: a randomized placebo-controlled trial. Hepatology 2004;40:1379–86. https://doi.org/10.1002/hep.20457.Search in Google Scholar PubMed
460. Silveira, MG, Torok, NJ, Gossard, AA, Keach, JC, Jorgensen, RA, Petz, JL, et al.. Minocycline in the treatment of patients with primary sclerosing cholangitis: results of a pilot study. Am J Gastroenterol 2009;104:83–8. https://doi.org/10.1038/ajg.2008.14.Search in Google Scholar PubMed
461. Shah, A, Macdonald, GA, Morrison, M, Holtmann, G. Targeting the gut microbiome as a treatment for primary sclerosing cholangitis: a conceptional framework. Am J Gastroenterol 2020;115:814–22. https://doi.org/10.14309/ajg.0000000000000604.Search in Google Scholar PubMed PubMed Central
462. Shah, A, Crawford, D, Burger, D, Martin, N, Walker, M, Talley, NJ, et al.. Effects of antibiotic therapy in primary sclerosing cholangitis with and without inflammatory bowel disease: a systematic review and meta-analysis. Semin Liver Dis 2019;39:432–41. https://doi.org/10.1055/s-0039-1688501.Search in Google Scholar PubMed
463. Wortelboer, K, Nieuwdorp, M, Herrema, H. Fecal microbiota transplantation beyond Clostridioides difficile infections. EBioMedicine 2019;44:716–29. https://doi.org/10.1016/j.ebiom.2019.05.066.Search in Google Scholar PubMed PubMed Central
464. Hvas, CL, Dahl Jørgensen, SM, Jørgensen, SP, Storgaard, M, Lemming, L, Hansen, MM, et al.. Fecal microbiota transplantation is superior to fidaxomicin for treatment of recurrent Clostridium difficile infection. Gastroenterology 2019;156:1324–32.e3. https://doi.org/10.1053/j.gastro.2018.12.019.Search in Google Scholar PubMed
465. Tan, P, Li, X, Shen, J, Feng, Q. Fecal microbiota transplantation for the treatment of inflammatory bowel disease: an update. Front Pharmacol 2020;11:574533. https://doi.org/10.3389/fphar.2020.574533.Search in Google Scholar PubMed PubMed Central
466. Tabibian, JH, O’Hara, SP, Trussoni, CE, Tietz, PS, Splinter, PL, Mounajjed, T, et al.. Absence of the intestinal microbiota exacerbates hepatobiliary disease in a murine model of primary sclerosing cholangitis. Hepatology 2016;63:185–96. https://doi.org/10.1002/hep.27927.Search in Google Scholar PubMed PubMed Central
467. Jiang, B, Yuan, G, Wu, J, Wu, Q, Li, L, Jiang, P. Prevotella copri ameliorates cholestasis and liver fibrosis in primary sclerosing cholangitis by enhancing the FXR signalling pathway. Biochim Biophys Acta, Mol Basis Dis 2022;1868:166320. https://doi.org/10.1016/j.bbadis.2021.166320.Search in Google Scholar PubMed
468. Liu, Y, Chen, K, Li, F, Gu, Z, Liu, Q, He, L, et al.. Probiotic Lactobacillus rhamnosus GG prevents liver fibrosis through inhibiting hepatic bile acid synthesis and enhancing bile acid excretion in mice. Hepatology 2020;71:2050–66. https://doi.org/10.1002/hep.30975.Search in Google Scholar PubMed PubMed Central
469. Roh, YS, Cho, A, Cha, YS, Oh, SH, Lim, CW, Kim, B. Lactobacillus aggravate bile duct ligation-induced liver inflammation and fibrosis in mice. Toxicol Res 2018;34:241–7. https://doi.org/10.5487/tr.2018.34.3.241.Search in Google Scholar
470. Shi, Y, Jiang, N, Li, M, Zeng, X, Tian, X. Mesenchymal stem cells and connective tissue diseases: from bench to bedside. J Transl Int Med 2023;11:30–45. https://doi.org/10.2478/jtim-2022-0028.Search in Google Scholar PubMed PubMed Central
471. Yang, Y, Zhao, RC, Zhang, F. Potential mesenchymal stem cell therapeutics for treating primary biliary cholangitis: advances, challenges, and perspectives. Front Cell Dev Biol 2022;10:933565. https://doi.org/10.3389/fcell.2022.933565.Search in Google Scholar PubMed PubMed Central
472. Wang, D, Zhang, H, Liang, J, Gu, Z, Ma, X, Huang, J, et al.. Effect of allogeneic bone marrow-derived mesenchymal stem cells transplantation in a polyI:C-induced primary biliary cirrhosis mouse model. Clin Exp Med 2011;11:25–32. https://doi.org/10.1007/s10238-010-0105-6.Search in Google Scholar PubMed
473. Lei, J, Chai, Y, Xiao, J, Hu, H, Liu, Z, Xiao, Y, et al.. Antifibrotic potential of bone marrow-derived mesenchymal stem cells in biliary atresia mice. Mol Med Rep 2018;18:3983–8. https://doi.org/10.3892/mmr.2018.9353.Search in Google Scholar PubMed
474. Cao, Y, Ji, C, Lu, L. Mesenchymal stem cell therapy for liver fibrosis/cirrhosis. Ann Transl Med 2020;8:562. https://doi.org/10.21037/atm.2020.02.119.Search in Google Scholar PubMed PubMed Central
475. Mancinelli, R, Franchitto, A, Gaudio, E, Onori, P, Glaser, S, Francis, H, et al.. After damage of large bile ducts by gamma-aminobutyric acid, small ducts replenish the biliary tree by amplification of calcium-dependent signaling and de novo acquisition of large cholangiocyte phenotypes. Am J Pathol 2010;176:1790–800. https://doi.org/10.2353/ajpath.2010.090677.Search in Google Scholar PubMed PubMed Central
476. Mancinelli, R, Franchitto, A, Glaser, S, Meng, F, Onori, P, Demorrow, S, et al.. GABA induces the differentiation of small into large cholangiocytes by activation of Ca(2+)/CaMK I-dependent adenylyl cyclase 8. Hepatology 2013;58:251–63. https://doi.org/10.1002/hep.26308.Search in Google Scholar PubMed PubMed Central
477. McDaniel, K, Meng, F, Wu, N, Sato, K, Venter, J, Bernuzzi, F, et al.. Forkhead box A2 regulates biliary heterogeneity and senescence during cholestatic liver injury in mice. Hepatology 2017;65:544–59. https://doi.org/10.1002/hep.28831.Search in Google Scholar PubMed PubMed Central
478. Sato, K, Meng, F, Venter, J, Giang, T, Glaser, S, Alpini, G. The role of the secretin/secretin receptor axis in inflammatory cholangiocyte communication via extracellular vesicles. Sci Rep 2017;7:11183. https://doi.org/10.1038/s41598-017-10694-3.Search in Google Scholar PubMed PubMed Central
479. Chaiyadet, S, Sotillo, J, Smout, M, Cantacessi, C, Jones, MK, Johnson, MS, et al.. Carcinogenic liver fluke secretes extracellular vesicles that promote cholangiocytes to adopt a tumorigenic phenotype. J Infect Dis 2015;212:1636–45. https://doi.org/10.1093/infdis/jiv291.Search in Google Scholar PubMed PubMed Central
480. Azar, J, Bahmad, HF, Daher, D, Moubarak, MM, Hadadeh, O, Monzer, A, et al.. The use of stem cell-derived organoids in disease modeling: an update. Int J Mol Sci 2021;22:7667. https://doi.org/10.3390/ijms22147667.Search in Google Scholar PubMed PubMed Central
481. Corrò, C, Novellasdemunt, L, Li, VSW. A brief history of organoids. Am J Physiol Cell Physiol 2020;319:C151–c65. https://doi.org/10.1152/ajpcell.00120.2020.Search in Google Scholar PubMed PubMed Central
482. Sidhaye, J, Knoblich, JA. Brain organoids: an ensemble of bioassays to investigate human neurodevelopment and disease. Cell Death Differ 2021;28:52–67. https://doi.org/10.1038/s41418-020-0566-4.Search in Google Scholar PubMed PubMed Central
483. Suezawa, T, Kanagaki, S, Moriguchi, K, Masui, A, Nakao, K, Toyomoto, M, et al.. Disease modeling of pulmonary fibrosis using human pluripotent stem cell-derived alveolar organoids. Stem Cell Rep 2021;16:2973–87. https://doi.org/10.1016/j.stemcr.2021.10.015.Search in Google Scholar PubMed PubMed Central
484. Picchio, V, Floris, E, Derevyanchuk, Y, Cozzolino, C, Messina, E, Pagano, F, et al.. Multicellular 3D models for the study of cardiac fibrosis. Int J Mol Sci 2022;23:11642. https://doi.org/10.3390/ijms231911642.Search in Google Scholar PubMed PubMed Central
485. Srivastava, V, Huycke, TR, Phong, KT, Gartner, ZJ. Organoid models for mammary gland dynamics and breast cancer. Curr Opin Cell Biol 2020;66:51–8. https://doi.org/10.1016/j.ceb.2020.05.003.Search in Google Scholar PubMed PubMed Central
486. Engevik, AC, Feng, R, Choi, E, White, S, Bertaux-Skeirik, N, Li, J, et al.. The development of spasmolytic polypeptide/TFF2-expressing metaplasia (SPEM) during gastric repair is absent in the aged stomach. Cell Mol Gastroenterol Hepatol 2016;2:605–24. https://doi.org/10.1016/j.jcmgh.2016.05.004.Search in Google Scholar PubMed PubMed Central
487. Estrada, HQ, Patel, S, Rabizadeh, S, Casero, D, Targan, SR, Barrett, RJ. Development of a personalized intestinal fibrosis model using human intestinal organoids derived from induced pluripotent stem cells. Inflamm Bowel Dis 2022;28:667–79. https://doi.org/10.1093/ibd/izab292.Search in Google Scholar PubMed PubMed Central
488. Bao, YL, Wang, L, Pan, HT, Zhang, TR, Chen, YH, Xu, SJ, et al.. Animal and organoid models of liver fibrosis. Front Physiol 2021;12:666138. https://doi.org/10.3389/fphys.2021.666138.Search in Google Scholar PubMed PubMed Central
489. Amarachintha, SP, Mourya, R, Ayabe, H, Yang, L, Luo, Z, Li, X, et al.. Biliary organoids uncover delayed epithelial development and barrier function in biliary atresia. Hepatology 2022;75:89–103. https://doi.org/10.1002/hep.32107.Search in Google Scholar PubMed PubMed Central
490. Liu, Y, Li, N, Zhu, Y. Pancreatic organoids: a frontier method for investigating pancreatic-related diseases. Int J Mol Sci 2023;24. https://doi.org/10.3390/ijms24044027.Search in Google Scholar PubMed PubMed Central
491. Gupta, N, Matsumoto, T, Hiratsuka, K, Garcia Saiz, E, Galichon, P, Miyoshi, T, et al.. Modeling injury and repair in kidney organoids reveals that homologous recombination governs tubular intrinsic repair. Sci Transl Med 2022;14:eabj4772. https://doi.org/10.1126/scitranslmed.abj4772.Search in Google Scholar PubMed PubMed Central
492. Sobral-Reyes, MF, Lemos, DR. Recapitulating human tissue damage, repair, and fibrosis with human pluripotent stem cell-derived organoids. Stem Cell 2020;38:318–29. https://doi.org/10.1002/stem.3131.Search in Google Scholar PubMed
493. Moran-Horowich, A, Lemos, DR. Methods for the study of renal fibrosis in human pluripotent stem cell-derived kidney organoids. Methods Mol Biol 2021;2299:435–45. https://doi.org/10.1007/978-1-0716-1382-5_29.Search in Google Scholar PubMed
494. Bowers, SLK, Meng, Q, Kuwabara, Y, Huo, J, Minerath, R, York, AJ, et al.. Col1a2-Deleted mice have defective type I collagen and secondary reactive cardiac fibrosis with altered hypertrophic dynamics. Cells 2023;12. https://doi.org/10.3390/cells12172174.Search in Google Scholar PubMed PubMed Central
495. Weiss, DJ. What is the need and why is it time for innovative models for understanding lung repair and regeneration? Front Pharmacol 2023;14:1130074. https://doi.org/10.3389/fphar.2023.1130074.Search in Google Scholar PubMed PubMed Central
496. Ma, L, Wu, Q, Tam, PK. The current proceedings of PSC-based liver fibrosis therapy. Stem Cell Rev Rep 2023;19:2155–65. https://doi.org/10.1007/s12015-023-10592-4.Search in Google Scholar PubMed
497. Sampaziotis, F, Muraro, D, Tysoe, OC, Sawiak, S, Beach, TE, Godfrey, EM, et al.. Cholangiocyte organoids can repair bile ducts after transplantation in the human liver. Science 2021;371:839–46. https://doi.org/10.1126/science.aaz6964.Search in Google Scholar PubMed PubMed Central
498. Cardinale, V, Wang, Y, Carpino, G, Cui, CB, Gatto, M, Rossi, M, et al.. Multipotent stem/progenitor cells in human biliary tree give rise to hepatocytes, cholangiocytes, and pancreatic islets. Hepatology 2011;54:2159–72. https://doi.org/10.1002/hep.24590.Search in Google Scholar PubMed
499. De Assuncao, TM, Sun, Y, Jalan-Sakrikar, N, Drinane, MC, Huang, BQ, Li, Y, et al.. Development and characterization of human-induced pluripotent stem cell-derived cholangiocytes. Lab Invest 2015;95:684–96. https://doi.org/10.1038/labinvest.2015.51.Search in Google Scholar PubMed PubMed Central
500. Shen, H. Core Concept: organoids have opened avenues into investigating numerous diseases. But how well do they mimic the real thing? Proc Natl Acad Sci U S A 2018;115:3507–9. https://doi.org/10.1073/pnas.1803647115.Search in Google Scholar PubMed PubMed Central
501. Ludwig, J, Barham, SS, LaRusso, NF, Elveback, LR, Wiesner, RH, McCall, JT. Morphologic features of chronic hepatitis associated with primary sclerosing cholangitis and chronic ulcerative colitis. Hepatology 1981;1:632–40. https://doi.org/10.1002/hep.1840010612.Search in Google Scholar PubMed
© 2024 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License.
Articles in the same Issue
- Frontmatter
- Editorial
- Hormone based therapy and crosstalk beyond hormones
- Reviews
- Dysregulated bile acid homeostasis: unveiling its role in metabolic diseases
- Beyond reproduction: unraveling the impact of sex hormones on cardiometabolic health
- Glucagon-like peptide (GLP)-1 regulation of lipid and lipoprotein metabolism
- Hepatic function of glucagon-like peptide-1 and its based diabetes drugs
- Biliary fibrosis is an important but neglected pathological feature in hepatobiliary disorders: from molecular mechanisms to clinical implications
Articles in the same Issue
- Frontmatter
- Editorial
- Hormone based therapy and crosstalk beyond hormones
- Reviews
- Dysregulated bile acid homeostasis: unveiling its role in metabolic diseases
- Beyond reproduction: unraveling the impact of sex hormones on cardiometabolic health
- Glucagon-like peptide (GLP)-1 regulation of lipid and lipoprotein metabolism
- Hepatic function of glucagon-like peptide-1 and its based diabetes drugs
- Biliary fibrosis is an important but neglected pathological feature in hepatobiliary disorders: from molecular mechanisms to clinical implications